

Overexpress of CD47 does not alter the stemness of MCF-7 breast cancer cells
Copyrights: Oanh Thi-Kieu Nguyen, Anh Nguyen-Tu Bui, Ngoc Bich Vu, Phuc Van Pham, 2016. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Background: CD47 is a transmembrane glycoprotein expressed on all cells in the body and particularly overexpressed on cancer cells and cancer stem cells of both hematologic and solid malignancies. In the immune system, CD47 acts as a “don’t eat me” signal, inhibiting phagocytosis by macrophages by interaction with signal regulatory protein α (SIRPα). In cancer, CD47 promotes tumor invasion and metastasis. This study aimed to evaluate the stemness of breast cancer cells when CD47 is overexpressed.
Methods: MCF-7 breast cancer cells were transfected with plasmid pcDNA3.4-CD47 containing the CD47 gene. The stemness of the transduced MCF7 cell population was evaluated by expression of CD44 and CD24 markers, anti-tumor drug resistance and mammosphere formation.
Results: Transfection of plasmid pcDNA3.4-CD47 significantly increased the expression of CD47 in MCF-7 cells. The overexpression of CD47 in transfected MCF-7 cells led to a significant increase in the CD44+CD24- population, but did not increase doxorubicin resistance of the cells or their capacity to form mammospheres.
Conclusion: CD47 overexpression enhances the CD44+CD24- phenotype of breast cancer cells as observed by an increase in the CD44+CD24- expressing population. However, these changes are insufficient to increase the stemness of breast cancer cells.
Introduction
According to World Health Organization (WHO), breast cancer is the most common cancer in women both in the developed and developing nations with nearly 1.7 million cases diagnosed in 2012 Ferlay et al., 2015. The main goal of cancer treatment is to cure or prolong the lifetime of patients and to ensure the best quality of life after treatments. By using a model in which human breast cancer cells were grown in immunocompromised mice, Al-Hajj at el., 2003 had found that only a minority of breast cancer cells had the ability to form new tumor Al-Hajj et al., 2003. Recently, much evidence was provided that leukemia and solid tumors maintenance and growth are sustained by a small proportion of cells exhibiting stem cell properties Nicolis, 2007Reya et al., 2001Visvader and Lindeman, 2008 which named tumorinitiating cells (TICs) or cancer stem cells (CSCs) Clarke et al., 2006. CSCs are resistant to many conventional cancer therapies, chemotherapy and radiotherapyso they can survive after treatments in spite of shrinkage of the tumor which results to tumor relapse. They are difficult to kill because they overexpress drug efflux pumps and have an increased capacity to active anti-apoptotic and pro-survival pathways as well as DNA repair Flemming, 2015. Actually, to ensure completely cure cancer, many new cancer therapies now are focused on the targeting CSCs. Recently, some studies showed that cancer stem cells could escape the imunne system by high expression of CD47 on their cell surface Cioffi et al., 2015Zhang et al., 2015.
Indeed, CD47 is expressed on almost all human cells as evaluated by mRNA expression and immunohistochemical staining. CD47 was first identified as an antigen expressed on ovarian carcinoma cells Mawby et al., 1994. CD47 is overexpressed in many solid tumors as well as hematologic malignancies Chao et al., 2010aChao et al., 2010bChao et al., 2012Jaiswal et al., 2009Majeti et al., 2009. CD47 is involved in the regulation of various activities of different immune cell types and can induce direct cancer cell death when it is crosslinked Oldenborg, 2004. In addition, CD47 plays an important role in angiogenesis, proliferation and cell adhesion Brown and Frazier, 2001Reinhold et al., 1995. The interaction between CD47 on T-cells and its receptor SIRP-α on dendritic cells (DCs) is part of a new regulatory pathway that may be involved in the maintenance of homeostasis by preventing the escalation of the inflammatory immune response Latour et al., 2001. Anti-CD47 antibody treatment not only enables macrophage-mediated phagocytosis of cancer cells but also can initiate an antitumor cytotoxic T-cell immune response Tseng et al., 2013. CD47 expression is elevated in human non-Hodgkin lymphoma (NHL); blocking anti-CD47 antibodies preferentially enabled phagocytosis of NHL cells and synergized with rituximab Chao et al., 2010a.
Therefore, we hypothesized that CD47 holds the important role in cancer stem cell formation. Therefore, this study aimed to over-express the CD47 in breast cancer cells and avaluate their stemness as breast cancer stem cells.
Material and methods
Cancer cell lines
Breast cancer cell lines (MCF-7 and MDA-M-231) were obtained from ATCC and VNBRC1; S6 cell line was established by our laboratory (Laboratory of Stem Cell Research and Application, University of Science, Vietnam National University, Ho Chi Minh city, Vietnam). All cell lines were thawed from nitrogen and cultured for several passages to confluency prior to use in experiments.
Cell culture
Cells were cultured in DMEM/F12 complete (i.e. DMEM/F12 with 10% fetal bovine serum and 1% antibiotic-mycotic; all reagents purchased from Sigma- Aldrich, St Louis, MO) in a 37°C, 5% CO2 incubator.
Cloning pcDNA3.4-CD47 plasmid
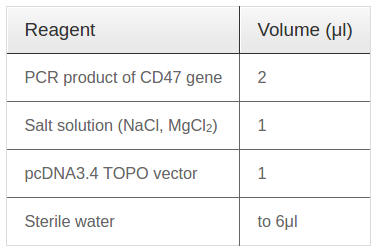
CD47 gene was isolated from cDNA of mRNA from S6 cells by PCR reaction with full-length primers under thermal cycle: denaturation 95°C / 5 minutes - 35 cycles (95°C / 20 seconds - 59°C / 30 seconds - 72°C / 1 min) - lasted 72°C / 10 minutes. Then, fresh PCR products were ligated with linear pcDNA3.4 vector (Life Technologies, USA) by TA cloning reaction and Topoisomerase enzyme which available in vector pcDNA3.4. The ingredient of ligation reaction followed Table 1 .
Transient transfection of DNA plasmid
DNA plasmids were transiently transfected by electroporation on a Multiporator®Electroporation system (Eppendorf, Hamburg, Germany) following the manufacturer’s instructions. In brief, MCF-7 cells were trypsinized, adjusted to a density of 0.5-1 x106cells/mL in 400 μL Hypoosmolar Solution (Eppendorf), and incubated with 20 μg/mL DNA plasmid. The electroporation was performed at 300 V in 40 μs. After electroporation, transfected cells were cultured in DMEM/F12 containing 10% FBS in a 37°C, 5% CO2 incubator. After 48 h of incubation, cells were analyzed for expression of CD47 and stemness-related genes, formation of spheres, and anti-tumor drug resistance.
Real-time RT-PCR
Approximately 5x106 cells were obtained for total RNA isolated using EasyBlue total RNA extraction kit (iNtRON, Korea) according to the manufacturer’s instructions. The concentration of total RNA were measured with a Biophotometer (Eppendorf). Realtime one step RT-PCR was carried out using a 2x qPCR SyGreen 1-Step Mix Lo-ROX (PCR BioSystems, London, UK). Relative quantification was estimated by the Livak method, 2-ΔΔct. Table 2

CD47 expression and the ratio of breast cancer cell populations were analyzed based on the percentage of cells expressing these markers: CD47, CD44 and CD24. CD44 is a marker of cancer stem cells Baumann and Krause, 2010 and CD24 is considered to be a marker for breast cancer cells Schabath et al., 2006. Cultured cells were trypsinized into single cells and diluted in 100 μL FACSflow solution at a density of 106cells/mL. Then cells were stained with CD47-FITC (Sigma-Aldrich), CD24-FITC (BD Biosciences, San Jose, CA) and CD44-APC antibodies (BD Biosciences). Cells were analyzed by flow cytometry on a FACSCalibur machine (BD Biosciences) using CellQuest Pro software (BD Biosciences).
Immunocytochemistry
Cells were cultured in 48-well plates at a cell density of 105 cells/well for 24 h before use in experiments.
Cells were fixed with 4% paraformaldehyde and washed three times with cold PBS. Fixed cells were incubated with anti-CD47 FITC-conjugated Ab (Sigma-Aldrich, Louis St, MO), anti-CD24 FITCconjugated Ab (BD Biosciences), and/or anti-CD44 PE-conjugated Ab (BD Biosciences). Cell nuclei were stained with Hoeschst 33442 (Sigma-Aldrich) and images were captured by a Carl Zeiss AxioLab A1 microscope (Carl Zeiss, Germany).
Sphere formation assay
Cells were plated at 103 cells/mL in serum-free DMEM/F12 medium supplemented with 10 ng/mL basic fibroblast growth factor (bFGF), 20 ng/mL epidermal growth factor (EGF), 5 ng/mL insulin (Santa Cruz Biotechnology, Santa Cruz, CA), and 0.4% bovine serum albumin (BSA; Sigma-Aldrich). Cells cultured under these conditions were capable of forming spherical clusters of cells (“spheres” or “mammospheres”). The number of spheres in each well was evaluated after 7 d of culture.
Anti-tumor drug assay
Non-transfected MCF-7 and CD47-transfected MCF-7 cells were plated at a density of 104 cells/well in Eplate 96 (ACEA Biosciences, San Diego) in DMEM/F12 supplemented with 10% FBS. After 24 h, cells were treated with 0, 1, 3, 6, or 9 μg/mL doxorubicin (Sigma- Aldrich) for 48 h. The cells were evaluated by an xCELLigence RTCA SP system for proliferation and drug resistance (IC50 index).
Statistical analysis
All experiments were performed in triplicate. Data were analyzed by Grapad Prism 6.0 software. The significant differences between mean values were assessed by the t-test and ANOVA, with P-value < 0.05 was considered to be significant.
Results
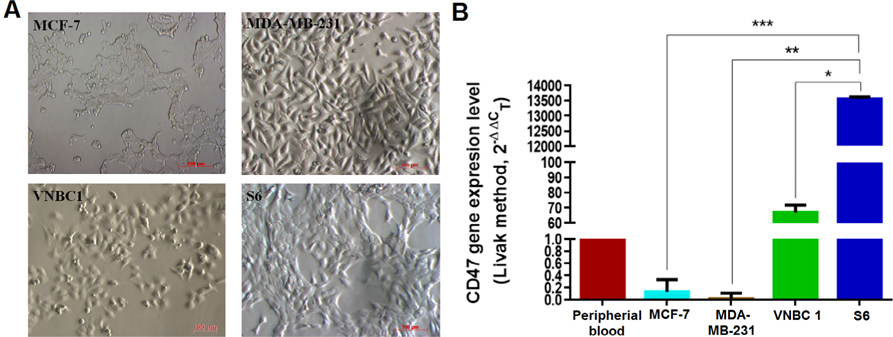
Morphology and CD47 gene expression in four breast cancer cell lines (MCF-7, MDA-MB-231, VNBRC1 and S6)
To determine CD47 expression in cancer cell lines, cells were cultured for real-time RT-PCR analysis using specific primers for CD47. When cultured as an adherent monolayer, MCF-7 cells exhibited an epithelial-like morphology (i.e. bean shape) while MDA-MB-231 cells exhibited a mesenchymal-like, spindle shape. VNBRC1 and S6 cells had an epitheliallike morphology. When observed under microscopy, MCF-7 cells were smaller in size than other three cancer cell lines ( Figure 1A ). CD47 gene expression was evaluated by real-time RT-PCR in three independent experiments. The results showed that when compared to CD47 mRNA expression by peripheral blood cells, the CD47 expression level by VNBRC1 cells was similar (average level), while expression by S6 cells was significantly higher (13556±67 fold) and by MDAMB- 231 cells was significantly lower (0.017±0.09 fold) ( Figure 1B ).
Expression of CD47, CD44, and CD24 cell populations in the four breast cancer cell lines
After trypsinization, expression of CD47, CD44 and CD24 markers was assessed on the four breast cancer cell lines by flow cytometry and immunocytochemistry. The results of flow cytometry showed that there are significant differences in CD47 expression by S6 and VNBRC1 cells, VNBRC1 and MDA-MB-231 cells, MDA-MB-231 and MCF-7 cells, and MCF-7 and S6 cells (97.38±0.35% vs. 93.6±0.42%, 93.6±0.42% vs. 87.91±1.16%, 87.91±1.16% vs. 74.86±1.34%, and 74.86±1.34% vs. 97.38±0.35%, respectively) (p < 0.05) ( Figure 2A ). Moreover, there were significant differences by flow in the percentage of the CD44+CD24- cell population for MDA-MB-231 vs.
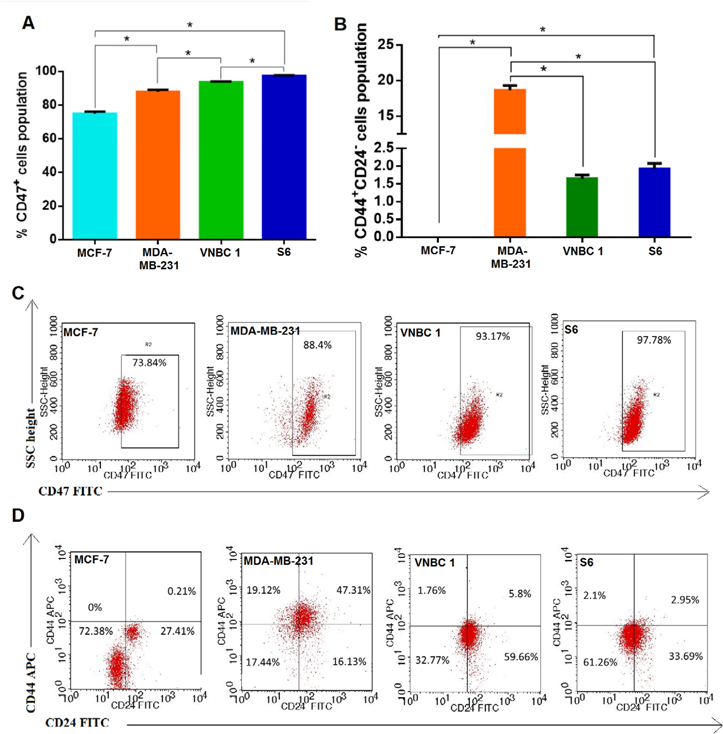
MCF-7, MDA-MB-231 vs VNBRC1, MDA-MB-231 vs S6, and S6 vs MCF-7 (18.62±0.64% vs. 0%, 18.62±0.64% vs. 1.64±0.1%, 18.62±0.64% vs. 1.92±0.16%, and 1.92±0.16% vs 0%, respectively (p < 0.05) ( Figure 2B ).
FACS results confirmed CD47 protein expression by the four cancer cell lines ( Figure 2C ) as well as their coexpression of CD44 and CD24. Further analysis showed that MCF-7 cells and S6 were predominantly CD44 positive and CD24 negative; the percent of cells that were CD44+CD24- in the MCF-7 cell line and the S6 cell line were 69.18±3.24 % and 60.15±0.97%, respectively. The percent of MDA-MB-231 cells with the CD44+CD24- phenotype was 47.52±0.48%, while the percent for VNBRC1 cells was 61.72±1.91%.
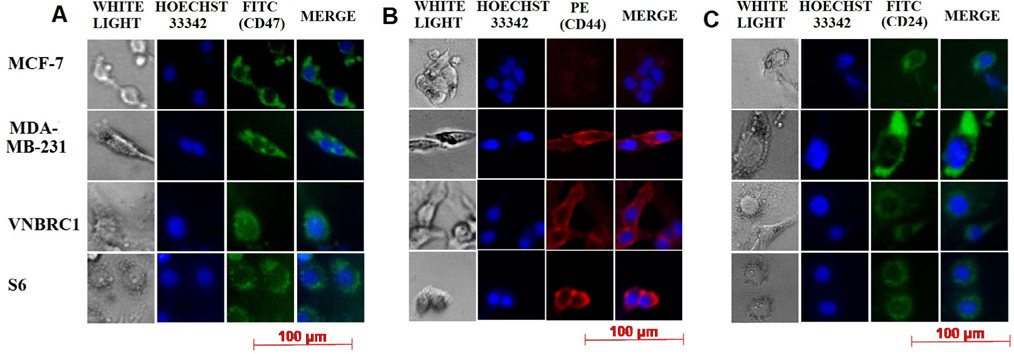
The results of immunocytochemistry showed that all cancer cell lines express CD47 ( Figure 3A ). All four cancer cell lines evaluated in this study were positive for CD24 with MDA-MB-231 showing the greatest expression. The MCF-7 cell line was negative for the CD44 marker and only weakly positive for CD24; these results are similar to flow cytometry results for MCF-7 cells (0% CD44+CD24- cell population). The MDA-MB-231 cell line is positive for all three markers with a strong FITC signal for the CD24 marker; notably, VNBRC1 and S6 cell lines are also positive for all three markers, with an especially strong signal for CD47 and CD44.
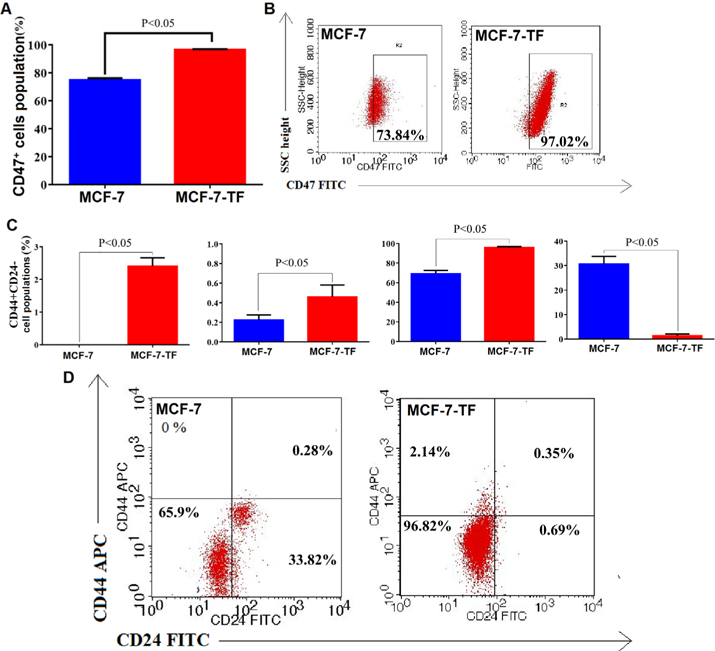
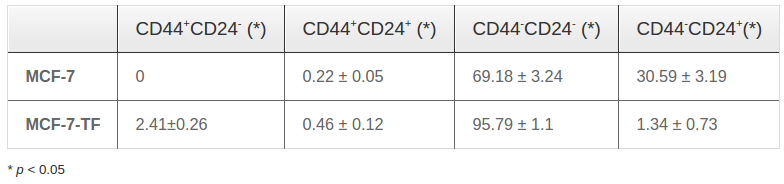
Expression of CD47 in CD47-transfected MCF-7 cells
Following pcDNA3.4-CD47 plasmid transfection, we performed protein quantification by flow cytometry. The results showed that the percent of CD47 positive cells increased significantly from 74.86±1.34% (nontransfected) to 96.43 ± 0.53% (transfected) (n=3 independent trials, p < 0.05) ( Figure 4A,B ). Flow cytometry results for CD44 and CD24 expression were shown in Table 1 . There were also significant differences in CD44 and CD24 expression by MCF-7 cells versus by transfected MCF-7 cells (called MCF-7- TF) ( Figure 4C,D ). Taken together, our results suggest that transfection efficiency can be enhanced via CD47, CD44 and CD24 markers on cell surfaces of MCF-7 cells. Table 3
Gene expression of several stemness-related genes
To investigate the increase of CD47 expression, total RNA was extracted to evaluate mRNA gene expression. In this study, we analyzed several genes related to CD47 and stemness in CD47-transfected MCF-7 cells (MCF-7-TF). We used normal MCF-7 cells as a negative control. This experiment was performed for 7 genes which profile is associated to stemness, and with GAPDH as an internal control. Real-time RTPCR were performed on 3 independent experiments and the relative quantitative results were analyzed by Livak method, 2-ΔΔC T.
As presented in Figure 5A , the expression of CD47 by MCF-7-TF (transfected) cells is higher than that for MCF-7 cells alone. The expression of OCT4, cMYC, SOX2, and NANOG by MCF-7-TF cells are higher than that for non-transfected MCF-7 cells. There are significant differences in CD47 expression between MCF-7 and MCF-7-TF cells (22.95±5.53 vs. 1), respectively. Moreover, there are similarities in the “stemness” gene expression of OCT4, cMYC, SOX2, NANOG, CD44, and CD24 in MCF-7 and MCF-7-TF cells (1.37±0.44 vs. 1; 1.33±1.09 vs. 1; 1.25±1.12 vs. 1; 2.14±1.74 vs 1, 0.85±0.51 vs. 1, and 1.17±0.13 vs 1, respectively) (p<0.05). These results demonstrate that upregulation of CD47 expression is not enough to alter the expression of several stemness genes in MCF- 7 cells, compared to pcDNA3.4-CD47 plasmid transfected MCF-7 cells.
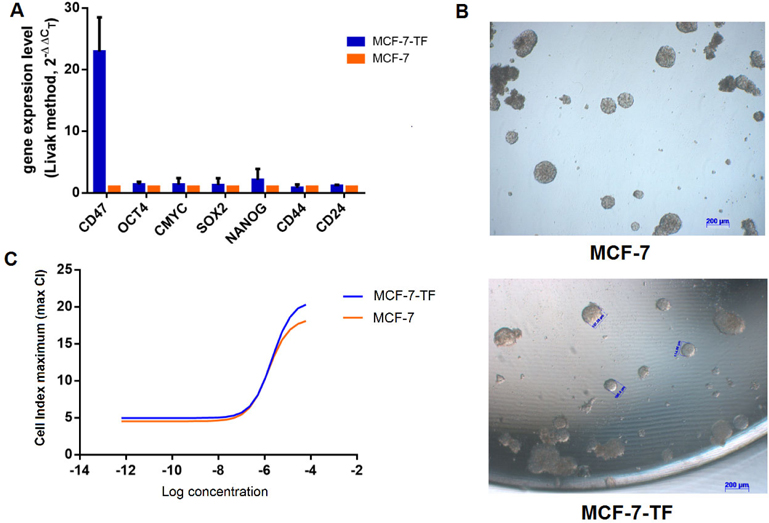
Sphere formation in serum-free medium
MCF-7 cells could not adhere onto the flask surface when cultured in serum-free medium nor could they form spheres ( Figure 5B ). There were similarities in the number of spheres in the MCF-7-TF group and MCF-7 group (95±12.6 and 98.67±26.4 spheres, respectively); however, there was no statistically significant difference (p=0.9062).
Drug resistance of MCF-7 after transfection with pcDNA3.4-CD47 plasmid
Doxorubicin had a suppressive effect on proliferation of breast cancer cells. In the absence of doxorubicin, proliferation rates were similar after 48 h in culture for non-transfected and CD47-transfected MCF-7 cell groups. There was some similarity in the IC50 index of non-transfected MCF-7 and MCF-7-TF (1.506 μg/mL and 2.1 μg/mL) ( Figure 5C ).
Discussion
From assessment of cell morphology, the results showed that three breast cancer cell lines (MCF-7, VNBRC1 and S6) are similar in that they have an epithelial-like shape. Conversely, the MDA-MB-231 cell line has a mesenchymal-like shape (spindle shape). The ratio of the CD44+CD24- population to the entire cell population for the VNBRC1 and S6 cell lines are similar to what has been described; some studies, too, have reported that less than 2% of the cell population have the CD44+CD24- phenotype Al-Hajj et al., 2003Appalaraju Jaggupilli, 2012Lapidot et al., 1994. CD47 expression in MCF-7 and MDA-MB-231 cells are similar to the published results of Manna et al. (2004); expression was monitored using an anti- CD47 antibody (BD Biosciences) Manna and Frazier, 2004.
There are significant differences in CD47 expression and the CD44+CD24- cell population of breast cancer cell lines. These differences may be caused by the origin and classification of the cancer cell lines. MCF-7 cells were derived from luminal epithelial cells while MDA-MB-231 cells were derived from basal epithelial cells so each cell line has a distinguishing gene profile Holliday and Speirs, 2011Kaur et al., 2015. CD47 mRNA and protein sequences are conserved among humans, mice and rats Chang and Huang, 2004. In this study, we showed the relationship between CD47 expression and the ratio of CD44+CD24- cells population in breast cancer cells. Taken together, the results demonstrated that S6 is a cell line with the highest CD47 expression; meanwhile, MCF-7showed the lowest CD47 expression among the four cell lines.
We used S6 cells for isolating the CD47 gene for subsequent cloning into the pcDNA3.4 vector to create a pcDNA3.4-CD47 plasmid. This plasmid was transfected into MCF-7 cells to induce upregulation of CD47 expression by MCF-7 cells. CD47 overexpression in MCF-7 cells was carried out by transfection of pcDNA3.4-CD47 plasmid into the cells. While electroporation is also a feasible procedure for gene delivery, DNA plasmid transfection was quite effective. In our study, the non-transfected MCF-7 cells did not express a CD44+CD24- population, similar to the results published by Sheridan et al., 2006. On the contrary, the transfected cells (MCF-7-TF) showed an increase in the CD44+CD24- cell population. We also found that CD47 expression is related to the ratio of the CD44+CD24- cell population; the changes in CD47 expression may cause changes in cell surface makers, thereby changing the phenotype of the cell population. However, these phenotypic changes could not modify cellular properties, such as drug resistance and mammosphere formation.
Gene expression of NANOG, OCT3/4, and SOX2 (stemness transcription factors) have been implicated in the progression of breast cancer Apostolou et al., 2012Apostolou et al., 2015. The oncogene cMYC is a transcription factor which regulates and initiates the immune regulator, CD47 Casey et al., 2016; overexpression of CD47 may impact the regulation by cMYC. Mammosphere formation in culture was used to study stem cell properties. It was noted that expression of SOX2 (but not OCT4 or NANOG) was induced; in fact, overexpression of SOX2 increased mammosphere formation and activation of breast cancer stem cells Leis et al., 2012.
According to particular properties of breast cancer stem cells, some breast cancer cells that are considered as breast cancer stem cells should exhibit stemness properties including both markers and biological activities. In our study, the cells either exhibited or promoted the CD44+CD24- phenotype, strong antitumor drug resistance, mammosphere formation, and tumors in NOD/SCID mice Jiao et al., 2016Pham et al., 2011Phan et al., 2016Saadin and White, 2013. Therefore, although overexpression of CD47 caused formation of the CD44+CD24- cell population, the cells did not promote the phenotypes of breast cancer stem cells. We propose that there are other signals besides CD47 which can alter breast cancer cells towards breast cancer stem cells.
Conclusion
The role of cancer stem cells in cancer recurrence and drug resistance is well-known. However, the relationship between cell surface markers and stemness properties of cancer stem cells is still unclear. This study showed that CD47 expression was related to formation of the CD44+CD24- cell population in MCF7 breast cancer cells. However, overexpression of CD47 could not change the anti-tumor drug resistance as well as impede mammosphere formation of these cells. Therefore, CD47 gene is not enough to induce MCF7 breast cancer cells to become breast cancer stem cells.
References
-
M.
Al-Hajj,
M.S.
Wicha,
A.
Benito-Hernandez,
S.J.
Morrison,
M.F.
Clarke.
Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S.
2003;
A100
:
3983-3988
.
-
P.
Apostolou,
M.
Toloudi,
M.
Chatziioannou,
E.
Ioannou,
I.
Papasotiriou.
Cancer stem cells stemness transcription factors expression correlates with breast cancer disease stage. Curr Stem Cell Res Ther.
2012;
7
:
415-419
.
-
P.
Apostolou,
M.
Toloudi,
I.
Papasotiriou.
Identification of genes involved in breast cancer and breast cancer stem cells. Breast Cancer (Dove Med Press).
2015;
7
:
183-191
.
-
E.E.
Appalaraju Jaggupilli.
Significance of CD44 and CD24 as Cancer Stem Cell Markers: An Enduring Ambiguity. Clinical and Developmental Immunology.
2012;
2012
:
11
.
-
M.
Baumann,
M.
Krause.
CD44: a cancer stem cellrelated biomarker with predictive potential for radiotherapy. Clin Cancer Res.
2010;
16
:
5091-5093
.
-
E.J.
Brown,
W.A.
Frazier.
Integrin-associated protein (CD47) and its ligands. Trends in Cell Biology.
2001;
11
:
130-135
.
-
S.C.
Casey,
L.
Tong,
Y.
Li,
R.
Do,
S.
Walz,
K.N.
Fitzgerald,
A.M.
Gouw,
V.
Baylot,
I.
Gutgemann,
M.
Eilers.
MYC regulates the antitumor immune response through CD47 and PD-L1. Science.
2016;
352
:
227-231
.
-
W.T.
Chang,
A.M.
Huang.
Alpha-Pal/NRF-1 regulates the promoter of the human integrin-associated protein/CD47 gene. J Biol Chem.
2004;
279
:
14542-14550
.
-
M.P.
Chao,
A.A.
Alizadeh,
C.
Tang,
J.H.
Myklebust,
B.
Varghese,
S.
Gill,
M.
Jan,
A.C.
Cha,
C.K.
Chan,
B.T.
Tan.
Anti-CD47 antibody synergizes with rituximab to promote phagocytosis and eradicate non-Hodgkin lymphoma. Cell.
2010a;
142
:
699-713
.
-
M.P.
Chao,
S.
Jaiswal,
R.
Weissman-Tsukamoto,
A.A.
Alizadeh,
A.J.
Gentles,
J.
Volkmer,
K.
Weiskopf,
S.B.
Willingham,
T.
Raveh,
C.Y.
Park.
Calreticulin is the dominant pro-phagocytic signal on multiple human cancers and is counterbalanced by CD47. Sci Transl Med.
2010b;
2
:
63ra94
.
-
M.P.
Chao,
I.L.
Weissman,
R.
Majeti.
The CD47- SIRPalpha pathway in cancer immune evasion and potential therapeutic implications. Curr Opin Immunol.
2012;
24
:
225-232
.
-
M.
Cioffi,
S.
Trabulo,
M.
Hidalgo,
E.
Costello,
W.
Greenhalf,
M.
Erkan,
J.
Kleeff,
B. Jr.
Sainz,
C.
Heeschen.
Inhibition of CD47 Effectively Targets Pancreatic Cancer Stem Cells via Dual Mechanisms. Clin Cancer Res.
2015;
21
:
2325-2337
.
-
M.F.
Clarke,
J.E.
Dick,
P.B.
Dirks,
C.J.
Eaves,
C.H.M.
Jamieson,
D.L.
Jones,
J.
Visvader,
I.L.
Weissman,
G.M.
Wahl.
Cancer Stem Cells-Perspectives on Current Status and Future Directions: AACR Workshop on Cancer Stem Cells. Cancer Research.
2006;
66
:
9339-9344
.
-
J.
Ferlay,
I.
Soerjomataram,
R.
Dikshit,
S.
Eser,
C.
Mathers,
M.
Rebelo,
D.M.
Parkin,
D.
Forman,
F.
Bray.
Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer.
2015;
136
:
E359-386
.
-
A.
Flemming.
Cancer stem cells: Targeting the root of cancer relapse. Nat Rev Drug Discov.
2015;
14
:
165
.
-
D.L.
Holliday,
V.
Speirs.
Choosing the right cell line for breast cancer research. Breast Cancer Res.
2011;
13
:
215
.
-
S.
Jaiswal,
C.H.
Jamieson,
W.W.
Pang,
C.Y.
Park,
M.P.
Chao,
R.
Majeti,
D.
Traver,
N.
van Rooijen,
I.L.
Weissman.
CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell.
2009;
138
:
271-285
.
-
X.
Jiao,
A.A.
Rizvanov,
M.
Cristofanilli,
R.R.
Miftakhova,
R.G.
Pestell.
Breast Cancer Stem Cell Isolation. Methods Mol Biol.
2016;
1406
:
121-135
.
-
S.
Kaur,
S.
Singh,
A.
Elkahloun,
W.
Wu,
T.
Song,
A.
Arora,
D.
Roberts.
Role of CD47 in triple negative breast cancer. The FASEB Journal.
2015;
29
.
-
T.
Lapidot,
C.
Sirard,
J.
Vormoor,
B.
Murdoch,
T.
Hoang,
J.
Caceres-Cortes,
M.
Minden,
B.
Paterson,
M.A.
Caligiuri,
J.E.
Dick.
A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature.
1994;
367
:
645-648
.
-
S.
Latour,
H.
Tanaka,
C.
Demeure,
V.
Mateo,
M.
Rubio,
E.J.
Brown,
C.
Maliszewski,
F.P.
Lindberg,
A.
Oldenborg,
A.
Ullrich.
Bidirectional negative regulation of human T and dendritic cells by CD47 and its cognate receptor signal-regulator protein-alpha: down-regulation of IL-12 responsiveness and inhibition of dendritic cell activation. J Immunol.
2001;
167
:
2547-2554
.
-
O.
Leis,
A.
Eguiara,
E.
Lopez-Arribillaga,
M.J.
Alberdi,
S.
Hernandez-Garcia,
K.
Elorriaga,
A.
Pandiella,
R.
Rezola,
A.G.
Martin.
Sox2 expression in breast tumours and activation in breast cancer stem cells. Oncogene.
2012;
31
:
1354-1365
.
-
R.
Majeti,
M.P.
Chao,
A.A.
Alizadeh,
W.W.
Pang,
S.
Jaiswal,
K.D. Jr.
Gibbs,
N.
van Rooijen,
I.L.
Weissman.
CD47 is an adverse prognostic factor and therapeutic antibody target on human acute myeloid leukemia stem cells. Cell.
2009;
138
:
286-299
.
-
P.P.
Manna,
W.A.
Frazier.
CD47 mediates killing of breast tumor cells via Gi-dependent inhibition of protein kinase A. Cancer Res.
2004;
64
:
1026-1036
.
-
W.J.
Mawby,
C.H.
Holmes,
D.J.
Anstee,
F.A.
Spring,
M.J.
Tanner.
Isolation and characterization of CD47 glycoprotein: a multispanning membrane protein which is the same as integrin-associated protein (IAP) and the ovarian tumour marker OA3. Biochem J 304(Pt.
1994;
2)
:
525-530
.
-
S.K.
Nicolis.
Cancer stem cells and “stemness” genes in neuro-oncology. Neurobiology of Disease.
2007;
25
:
217-229
.
-
P.A.
Oldenborg.
Role of CD47 in erythroid cells and in autoimmunity. Leuk Lymphoma.
2004;
45
:
1319-1327
.
-
P.V.
Pham,
N.L.
Phan,
N.T.
Nguyen,
N.H.
Truong,
T.T.
Duong,
D.V.
Le,
K.D.
Truong,
N.K.
Phan.
Differentiation of breast cancer stem cells by knockdown of CD44: promising differentiation therapy. J Transl Med.
2011;
9
:
209
.
-
N.L.
Phan,
N.V.
Trinh,
P.V.
Pham.
Low concentrations of 5-aza-2’-deoxycytidine induce breast cancer stem cell differentiation by triggering tumor suppressor gene expression. Onco Targets Ther.
2016;
9
:
49-59
.
-
M.I.
Reinhold,
F.P.
Lindberg,
D.
Plas,
S.
Reynolds,
M.G.
Peters,
E.J.
Brown.
In vivo expression of alternatively spliced forms of integrin-associated protein (CD47). J Cell Sci 108.
1995;
(Pt 11)
:
3419-3425
.
-
T.
Reya,
S.J.
Morrison,
M.F.
Clarke,
I.L.
Weissman.
Stem cells, cancer, and cancer stem cells. Nature.
2001;
414
:
105-111
.
-
K.
Saadin,
I.M.
White.
Breast cancer stem cell enrichment and isolation by mammosphere culture and its potential diagnostic applications. Expert Rev Mol Diagn.
2013;
13
:
49-60
.
-
H.
Schabath,
S.
Runz,
S.
Joumaa,
P.
Altevogt.
CD24 affects CXCR4 function in pre-B lymphocytes and breast carcinoma cells. J Cell Sci.
2006;
119
:
314-325
.
-
C.
Sheridan,
H.
Kishimoto,
R.K.
Fuchs,
S.
Mehrotra,
P.
Bhat- Nakshatri,
C.H.
Turner,
R. Jr.
Goulet,
S.
Badve,
H.
Nakshatri.
CD44+/CD24- breast cancer cells exhibit enhanced invasive properties: an early step necessary for metastasis. Breast Cancer Res.
2006;
8
:
R59
.
-
D.
Tseng,
J.P.
Volkmer,
S.B.
Willingham,
H.
Contreras-Trujillo,
J.W.
Fathman,
N.B.
Fernhoff,
J.
Seita,
M.A.
Inlay,
K.
Weiskopf,
M.
Miyanishi.
Anti-CD47 antibodymediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response. Proc Natl Acad Sci U S A.
2013;
110
:
11103-11108
.
-
J.E.
Visvader,
G.J.
Lindeman.
Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer.
2008;
8
:
755-768
.
-
H.
Zhang,
H.
Lu,
L.
Xiang,
J.W.
Bullen,
C.
Zhang,
D.
Samanta,
D.M.
Gilkes,
J.
He,
G.L.
Semenza.
HIF-1 regulates CD47 expression in breast cancer cells to promote evasion of phagocytosis and maintenance of cancer stem cells. Proc Natl Acad Sci U S A.
2015;
112
:
28
.
Comments
Downloads
Article Details
Volume & Issue : Vol 3 No 09 (2016)
Page No.: 826-835
Published on: 2016-09-29
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 10484 times
- Download PDF downloaded - 2132 times
- View Article downloaded - 18 times

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}