

Copyrights: Rintu Das, Pramathadhip Paul, Kaustab Mukherjee, Shinjini Mitra, Umesh Prasad Singh, Ena Ray Banerjee, 2015. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Fruit of date palm (Phoenix sylvestrisL.) is edible and used as an anti-geriatric, anti-oxidant ethnomedicine. In this study, three different types ofdateextracts, methanolic, acidic ethanolic and basic ethanolicwere evaluated for their putative in vitro scavenging effects on reactive oxygen species (ROS), where scavenging of hydroxyl radicals (basic ethanolic>acidic ethanolic>methanolic), superoxide radicals (acidic ethanolic>basic ethanolic>methanolic), DPPH radicals (acidic ethanolic>methanolic>basic ethanolic), nitric oxide (NO) (methanolic>acidic ethanolic>basic ethanolic) and inhibition of lipid peroxidation (basic ethanolic>acidic ethanolic> methanolic) were found to occur in a dose dependent manner. Their flavonoid and phenolic contents proved to be the source of this potent free radical scavenging activity and indicated a direct correlation with their total anti-oxidant capacity. On human embryonic kidney cell line (HEK) and murine RAW macrophages, bacterial lipopolysaccharide (LPS) induced inflammation, the date extracts applied therapeutically, inhibit intracellular oxidative stress significantly. This reinstatement of cellular homeostasis presumably occurs via mitochondrial pathways.
Introduction
One of the most primitive and yet effective mechanisms the mammalian physiology adopts in an exigent situation, which although predominantly nonspecific is remarkably fast and conclusive for the next phases of immune response or structural tissue remodeling, is inflammation. Inflammatory response occurs in a phasewise manner Crunkhorn and Meacock, 1971. Inflammation is the principal and sometimes critical initiator of most disorders. A crosstalk between the structural and immune cells causes destruction on the one hand and healing or reconstruction on the other. In diseases such as asthma, rheumatoid arthritis, psoriasis, multiple sclerosis, obesity and inflammatory bowel disease Winter et al., 1962, the first phase of disease onset, followed by establishment, development, maintenance and exacerbations, leads to a completion of various steps that are characteristic of the particular inflammatory disease. Although specific characteristics of inflammatory response in each disease and their site ofoccurrence may vary, a universal feature governing this stepwise phenomenon is the complex interplay amongstthe various cell subsets of the inflammatory cascade and tissue resident cells, and the network of signaling governing the two Vinegar et al., 1969Winter et al., 1962, that affect various target tissues Cortran, 2010.
Unmet needs in medicine and unknown phenomena prevailing in the mechanism of disease onset remain Mohan, 2010. In traditional medicine, that has provided solution to prevailing health issues at a global level, medicinal plants continue to provide valuable therapeutic agents. To avoid and mitigate various side effects and complications of modern medicine, and to address unmet needs of diseases, especially in the context of emerging complex etiopathophysiological pathways, traditional medicine is gaining importance and is now being studied systematically using biotechnological tools, to find the scientific basis of their therapeutic actions.
Phytochemicals from fruits and other edible plant parts have been shown to possess significant antioxidant properties that may be associated with lower incidence and lower mortality rates of degenerative diseases in human. Different biological properties, antioxidant capacities and radicalscavenging activities of various herbal extracts have been widely demonstrated, using in vitro techniques and in vivo models by
different groups of researchers Amann et al., 1995Chatpalliwar VA, 2002. The antiproliferative and antiinflammatory activities of these herbal extracts have been documented in human oral, breast, colon, cervical, and prostate cancer cell lines as well as in preclinical animal models by attenuating certain inflammatory intermediates, including nitric oxide, NFkB, and TNFα Miles and Miles, 1952Whittle, 1964.
This study was designed to identify antiinflammatory potential in extracts from datewhich is a wellknown ethno-medicine with high nutritive value. Among its contents, large amounts of several phenolic and nonphenolic compounds and other uncharacterized moieties may contribute to its use as a food supplement, a functional food or a nutraceutical substance with prophylactic and therapeutic functions in oxidative inflammatory diseases. Antioxidative and anti inflammatory activities of three types of date palm extracts, namely methanolic, basic ethanolic and acidic ethanolic, have been assessed, compared and validated, using both murine and human cell lines, in which inflammation has been induced by E. coli LPS, a potent pro inflammatory agent. In addition, intracellular cell organelle specific targeting of this extract has also been assessed in order to seek information regarding its mode of action in modulating the inflammatory cascade in a biological system, which is the most critical part for any drug discovery programme addressing inflammatory disorders.
Material and Methods
Extraction of date
Mesocarp tissue of date fruit was macerated and treated with 50% watermethanol solution at 40C. This was then divided into two parts: part A was evaporated to dryness, the remaining part was rinsed with water, dried at 500C, alkaline hydrolyzed with sodium hydroxide (0.1M) and filtered. The filtrate was acidified at pH 2.0, extracted with ethyl acetate and rinsed with 1% sodium bicarbonate and water. The organic layer was evaporated to dryness and then dissolved in water, to give part B. The residue after filtering out part B was alkaline hydrolyzed with 2M sodium hydroxide and filtered. The filtrate was further acidified at pH 2, extracted with ethyl acetate, rinsed with 1% sodium bicarbonate and water, and the organic layer evaporated to dryness (part C) ( Figure 1 ).
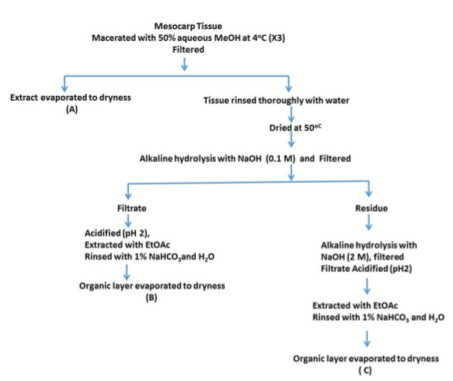
Reagents
Chemicals, such as ethylenediamine tetra acetic acid (EDTA), trichloroacetic acid (TCA), butanol, ammonium molybdate, and sodium dodecyl sulphate, benzoic acid, sodium phosphate, DMSO were purchased from E. Merck (India) Limited.1,1-Diphenyl-2-picrylhydrazyl and malondialdehyde, potassium ferricyanite, thiobarbituric acid (TBA) were procured from Sigma, USA. Nbutanol, ferrous sulphate, ferricchloride, Folins reagent, riboflavin, naphthylethylene diaminedihydrochloride, sulphanilamide inphosphoric acid, sodium bicarbonate, sodium hydroxide, and potassium hydroxide were purchased from Sisco Research Laboratories PVT. Ltd India. Nitroblue tetrazolium (NBT), MTT reagent [(4,5 dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide], DMEM media were purchased from Himedia, India. Fetal bovine serum (FBS) was purchased from Gibco. DCFHDA and DHR 123 were purchased from Invitrogen. All other reagents were of analytical grade.
DPPH radicalscavenging activity
The antioxidant activity of the extracts was measured on the basis of the scavenging activity of the stable 1,1-diphenyl-2-picrylhydrazyl (DPPH) free radical Braca et al., 2001. Aqueous extract was added to a 0.004% Methanol solution of DPPH on a 96 well ELISA plate.
Absorbance at 517 nm was determined after 30 min, and the percent inhibition activity was calculated.
Assay of superoxide radical O2°scavenging activity
The method used by Martinez et al. Martinez et al., 2001 for determination of the superoxide dismutase was studied in the riboflavinlight
nitrobluetetrazolium (NBT) system Beauchamp and Fridovich, 1971. Each 0.1 ml of reaction mixture contained 50 mM phosphate buffer (pH 7.8), 13 mM methionine, 2 µμM riboflavin, 100 µμM EDTA, NBT (75 µμM) and various doses of sample solution. The production of blue formazan was followed by monitoring the increase in absorbance at 560 nm after 15 min of illumination from a fluorescent lamp.
Assay of hydroxyl radical (OH)-scavenging activity
The assay was based on the benzoic acid hydroxylation method Chung et al., 1997. Hydroxyl radicals were generated by direct addition of iron(II) salts to a reaction mixture containing phosphate buffer. In a 24 well plate, 0.15 ml of sodium benzoate (10 mM) and 0.15 ml of FeSO4.7H2O (10 mM) and EDTA (10 mM) were added. Then the sample solution and a phosphate buffer (pH 7.4, 0.1 M) were added to give a total
volume of 1.6 ml. Finally, 0.15 ml of an H2O2 solution (10 mM) was added. The reaction mixture was then incubated at 370C for2 h. After that, the fluorescence was measured at 407nm emission (Em) and excitation (Ex) at 305 nm. Measurement of spectrofluorometric changes has been used to detect the damage by the hydroxyl radical.
Lipid peroxidation assay
A modified thiobarbituric acidreactive species (TBARS) assay Ohkawa et al., 1979 was used to measure the lipid peroxide formed, using egg yolk homogenates as lipidrich media Ruberto et al., 2000, where lipid peroxidation was induced by FeSO4.Malondialdehyde (MDA), produced by the oxidation of polyunsaturated fatty acids, reacts with two molecules of thiobarbituric acid (TBA), yielding a pinkish red chromogen with an absorbance maximum at 532 nm, which was measured using a 96 well ELISA plate reader. Percentage inhibition of lipid peroxidation by different concentrations of the extract was calculated.
Nitric oxide (NO) scavenging activity
Nitric oxide was generated from spontaneous decomposition of the Sodium nitropruside (20mM) in phosphate buffer (pH 7.4) which interacts with oxygen molecule to produce nitrite ions, which can be measured by the Griess reactions. The nitric oxide scavenging activity of the date extracts, collected by three different methods was determined by Shirwaikar et al (2006) with a slight modification. Briefly a stock solution of each extract was prepared to contain 1 mg/ml. Different amounts of the stock solution were then transferred to different test tubes and volume was adjusted to 1ml by adding double distilled water. 0.2 ml of sodium nitroprusside (20mM) in PBS (pH 7.4) and 1.8 ml of PBS solution was added and incubated at 37°C for 3 h. 1 ml of each solution was taken and diluted with 1 ml of Griess reagent [1% sulfanilamide, 5% H3PO4and 0.1% N(1naphthyl)ethylenediamine]. Similarly, a blank was prepared containing the equivalent amount of reagents (only the sodium nitroprusside and PBS), but without the extract. The absorbances of these solutions were measured at 540 nm against the corresponding blank solution. Ascorbic acid (100 µμg/ml) was used as the positive control. The percentage inhibition of nitric oxide was calculated.
Determination of total antioxidant capacity
The assay is based on the reduction of Mo (VI) to Mo (V) by the extract and subsequent formation of a green phosphate/Mo(V) complex at acid pH Prieto et al., 1999. Each well of a 96 well ELISA plate containing extract and reagent solution (0.6 M sulfuric acid, 28 mM sodium phosphate and 4 mM ammonium molyb date) were incubated at 950C for 90 min. After the mixture had cooled to room temperature, the absorbance of each solution was measured at 695 nm against a blank. The antioxidant capacity was expressed as ascorbic acid equivalent (AAE).
Determination of reducing power
The reducing power of date extracts was determined according to the method Salah et al., 1995 where different concentrations of extracts were mixed with phosphate buffer and potassium ferricyanide. The mixture was incubated at 50°C for20 min. 2.5 ml of trichloroacetic acid was added to the mixture. The upper layer of solution (2.5 ml) was mixed with distilled water (2.5 ml) and FeCl3, and the absorbance was
measured at 700 nm. Increased absorbance of the reaction mixture indicated increased reducing power. Ascorbic acid was used as positive control.
Determination of total flavonoid content
Total flavonoid content was determined using aluminium chloride (AlCl3) according to a known method, using Fisetin as a standard. The date extracts (0.1 ml) were added to 0.3 ml distilled water followed by 5% NaNO2 (0.03 ml). After 5 min at 25°C, AlCl3 (0.03 ml, 10%) was added. After further 5 min, the reaction mix ture was treated with 0.2 ml of 1 mM NaOH. Finally, the reaction mixture was diluted to 1 ml with water and the absorbance was measured at 510 nm. The results were expressed as mg Fisetin/g date extract.
Determination of total phenolic content
The total phenolic content of the date extracts was determined using the FolinCiocalteu reagent. The reaction mixture contained: 200 µμl of diluted extract, 800 µμl of freshly prepared diluted FolinCiocalteu reagent and 2 ml of 7.5% sodium carbonate. The final mixture was diluted to 7 ml with deionized water. Mixtures were kept in dark at ambient conditions for2 h to complete the reaction. The absorbance at 765 nm was measured. Gallic acid was used as standard and the results were expressed as mg gallic acid (GAE)/g of the date extract.
UV based spectrophotometric Analysis:
The samples of various extracts was analysed using UVVis Spectrophotometer. About 40 mg of the airdried samples was dissolved in 1 ml methanol, dilutedto 1 mg/ml solution in different volumetric flasks and then applied on the spectrophotometer and scanned through the UV and visible region. The herbal functional groups were determined by analyzing the peaks.
Cell culture
RAW 267.4 murine macrophage cell line and HEK 293 cell line were obtained from NCCS, Pune, India. The cells were grown in DMEM medium containing 5% inactivated fetal bovine serum, penicillin (100 U/mL), and streptomycin (20 µμg/mL) and kept at 37°C in a T25 tissue culture flasks. Cell were grown to confluence in a humidified atmosphere containing 5% CO2. Cell viability using MTT assay To test the cytotoxicity, 5×104 cells/well were seeded in a 96well plate and incubated for24 h with different concentrations of the date extract. The cells were washed, and each well was filled with 100 µμL of medium and 10 µμL of a tetrazolium salt, MTT. The plate was incubated for various time periods, and the absorbance was measured at 540 nm. The percentage of viable cells was calculated using the absorbance of the control cells without extract as 100%. The assay was performed in triplicate twice.
Proteomic analyses of extracts
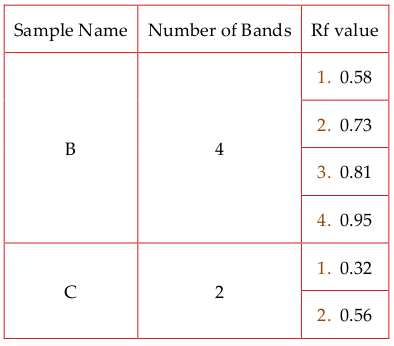
HPLC analyses: Crude methanolic A, basic ethanolic B & acidic ethanolic C were filtered by 0.22 µμm syringe filter (Milipore, Germany), and then the extracts were analysed with RPHPLC by using Waters 515 System with C18 column as stationary phase. The mobile phase was Water (HPLC grade) and 90% acetonitrile. Sample volume was 100 µμl. Flow rate was 0.5ml/min.TLC analyses: Methanolic extract (A), Acidic Ethanolic extract (B) and Basic ethanolic extract (C) were applied on silica gel coated TLC plates (Milipore, Germany) by using capillary tubes and developed in a TLC chamber using mobile phase methanol:water:chloroform (2.5:0.2:7.3 v/v). The developed TLC plates were air dried and observed under ultra violet light UV at both 254 nm and 366 nm. TLC plates were sprayed with Libermann’s solution, heated at 100–105°C and visualized under daylight. The movement of the analyte was expressed by its retention factor (Rf).
Methods
Taking 0% inhibition in the mixture without plant extract, regression equations were prepared from the concentrations of the three date extracts which were collected by three different methods and percentage inhibition of free radical formation/prevention in different systems of assay were calculated, viz. DPPH assay, superoxide radical scavenging assay, hydroxyl radicals scavenging assay, lipid peroxidation assay and nitric oxide radicals scavenging assay. IC50 values (concentration of sample required to scavenge 50% of available free radicals or to prevent lipid peroxidation by 50%) were calculated from these regression equations. IC50 value is inversely related to the activity of the extracts.
Assay of hydroxyl radical (OH)scavenging activity
Superoxide radical was measured by the NBT reduction assay. Each well of a 96wells plate was seeded with RAW 264.7 macrophages suspension containing .5×104 cells/ml. The treatment of cells proceeded as described previously. After incubation, 40 µμL of a NBT solution at 1 mg/mL was added to the medium and incubated at 37°C, for 1 h. Then, the incubation medium was removed and cells were lysed with DMSO:2 M NaOH (1:1). The absorbance of reduced NBT, formazan, was measured at 620 nm, in a microplate reader (Multiskan ASCENT Thermo®).
NO estimation
In culture, the NO released by the macrophages into the medium is converted to several nitrogen derivatives, from which only nitrite is stable, being easily measured by Griess reagent (1.0% sulphanilamide and 0.1% N(1)naphthylethylenediamine in 5% phosphoric acid). After incubation, 100 µμL of culture medium supernatant was mixed with the same volume of Griess reagent, during 10 min, at room temperature. The nitrite produced was determined by measuring the optical density at 540 nm, in a microplate reader (Shimadzu).
ROS measurement
Intracellular formation of ROS was assessed by using oxidation sensitive dye DCFHDA as a substrate Wijesinghe et al., 2011. RAW macrophage cells were seeded in 24well black plate at a concentration of 5x104 cells/ml. Cells were treated with 1 µμg/ml of E.coli LPS and then therapeutically various concentrations of date extracts were added on to it and incubated for 6 h. Negative control cells, i.e. only LPS treated as well as cells without any treatment were incubated for the same time period and then washed in PBS and after the addition of DCFDA (5 µμg/mL) incubate for 30 min at 37oC in dark. Nonfluorescent DCFHDA dye, that freely penetrate into cells get hydrolyzed by intracellular esterase to 207dichlorofluorescin (DCFH), and is trapped inside the cells. The formation of 207dichlorofluorescin (DCF) due to oxidation of DCFH in the presence of ROS was read after 30 min at an excitation wavelength of 485 nm and emission wavelength of 525 nm using a spectrofluorometer. Percent scavenging power of hydroxyl, superoxide and peroxide radicals was computed taking that by Ascorbic acid as 100%.
Measurement of mitochondrial membrane potential (MMP)
Mitochondrial membrane potential was monitored by the fluorescent dye, Rhodamine123. It is a cell permeable cationic dye that preferentially enters into mito chondria based on highly negative mitochondrial membrane potential (Wm). Depolarization of MMP results in the loss of Rhodamine 123 from the mitochondria and a decrease in intracellular fluorescence intensity. After the addition of 1 µμg/ml E.coli LPS, var
ious concentrations of date extracts were added to RAW macrophages cell line and incubated for 6h in 37oC. After incubation, the cells werewashed twice in cold PBS, then Rhodamine 123 (10 µμM) was added and incubated for 30 min at 37oC in dark. Fluorescence was measured by spectrofluorometer with an excitation wavelength of 485 nm and emission wavelength of 525 nm.
TLC of Khejur Extracts
Methanolic extract (A), Acidic Ethanolic extract (B) and Basic ethanolic extract (C) were applied on silicagel coated TLC plates (Milipore, Germany) by using capillary tubes and developed in a TLC chamber using mobile phase methanol:water:chloroform (2.5: 0.2:7.3 v/v). The developed TLC plates were air dried and observed under ultra violet light UV at both 254 nm and 366 nm. TLC plates were sprayed with Libermann’s solution, heated at 100–105°C and visualized under daylight. The movement of the analyte was expressed by its retention factor (Rf).
HPLC Analysis
Crude methanolic A, basic ethanolic B & acidic ethanolic C were filtered by 0.22 µμm syringe filter (Milipore, Germany), and then the extracts were analysed with RPHPLC by using Waters 515 System with C18 column as stationary phase. The mobile phase was Water (HPLC grade) and 90% acetonitrile. Sample volume was 100 µμl. Flow rate was 0.5 mL/min.
Statistical analysis
Statistical differences among samples were tested by Student t test. A Pvalue less than 0.05 or 0.01 (as applicable visàvis the assay performed) was considered statistically significant.
Results
Taking 0% inhibition in the mixture without plant extract, regression equations were prepared from the concentrations of the three date extracts which were collected by three different methods and percentage inhibition of free radical formation/prevention in different systems of assay were calculated, viz. DPPH assay, superoxide radical scavenging assay, hydroxyl radicals scavenging assay, lipid peroxidation assay and nitric oxide radicals scavenging assay. IC50 values (concentration of sample required to scavenge 50% of available free radicals or to prevent lipid peroxidation by 50%) were calculated from these regression equations. IC50 value is inversely related to the activity of the extracts ( Figure 2 ).
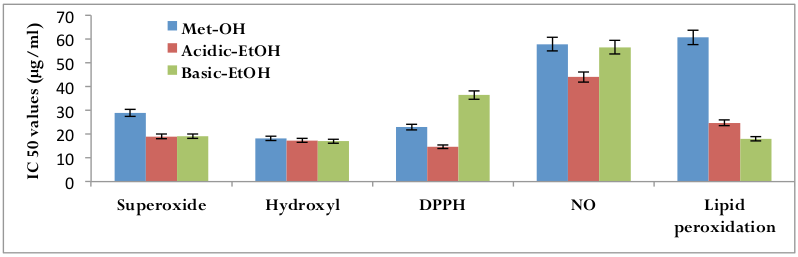
Various concentrations of date extracts, collected by three distinct methods, required to scavenge 50% free radical or to prevent lipid peroxidation by 50%. IC50 value is inversely related to the activity of the extracts. Both acidic and ethanolic date extracts are more active in scavenging the superoxide ions in anin vitro assay system than the methanolic date extracts, where as there is no significant variation in any of the three extracts in scavenging the hydroxyl radical. The DPPH radical scavenging and nitric oxide scavenging activitiesare greater in acidic ethanolic extracts in comparison to the methanolic extracts and basic ethanolic extracts respectively. Interestingly prevention of lipid peroxidation activity is highest in basic ethanolic extracts than the acidic ethanolic extracts and methanolic extracts. So, the ability to scavenge various free radicals or to prevent lipid peroxidation is different in all three extracts and it exerts the activity in a dose de-‐pendent manner.
DPPH radicalscavenging activity
Antioxidants, on interaction with DPPH transfer an electron (hydrogen atom) to DPPH, neutralizing its free radical character Naik et al., 2003. The colour changes from purple to yellow and its absorbance at wavelength 517 decreases. Various concentrations of date extracts, collected by three distinct methods, quenched DPPH free radical in a dosedependent manner [r2 =0.909 (p<0.01) for methanolic extract; r2 = 0.933 (p<0.001) for acidic ethanolic extract; r2 =0.930 (p<0.001) for basic ethanolic extract]. IC50 values were 22.91 μμg/ml for methanolic extract, 14.61 μμg/ml for acidic ethanolic extract and 36.44 μμg/ml for basic ethanolic extract. DPPH assay shows that, in this system, the radicalscavenging activities of the three varieties of date extracts are in the order acidic ethanolic extract>methanolic extract>basic ethanolic extract ( Figure 3 ).

Assay of hydroxylradical (OH)-scavenging activity
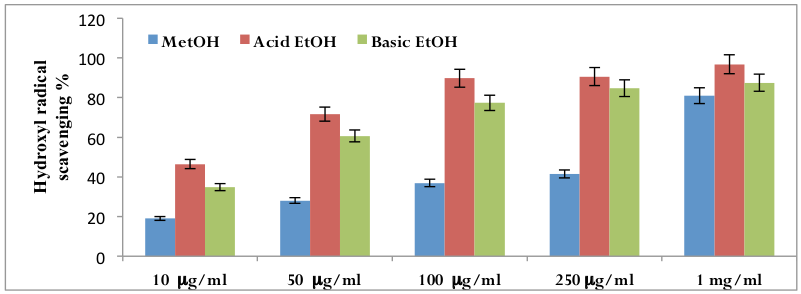
By the addition of iron (II) salts to a phosphate buffer containing reaction mixture, hydroxyl radicals can be generated Gutteridge, 1983. Benzoate, weakly fluorescent, after monohydroxylation forms highly fluorescent hydroxybenzoates Gutteridge, 1987. Measurement of this spectrofluorometric change has been used to detect damage by hydroxyl radical. Date ex-‐tracts collected by three distinctly separate methods was found to be a powerful scavenger of hydroxylradicals. There is a linear correlation between concentration of extract and OHscavenging activity [r2 =0.832 (p<0.01) for methanolic extracts of date; r2 =0.835 (p< 0.01) for acidic ethanolic extracts of date; r2 = 0.882 (p<0.01) for basic ethanolic extracts of date]. IC50 values are 18.20 μμg/ml for methanolic date extracts, 17.30 μμg/ml for acidic ethanolic date extracts and 17.00 μμg/ml for basic ethanolic extracts. Highest hydroxyl radicalscavenging activity was found in the date extract collected by using basic ethanolic methods. The hydroxyl radical scavenging properties of date extracts are: basic ethanolic extracts> acidic ethanolic extracts > methanolic extracts ( Figure 4 ).
Superoxide radical O2° scavenging activity
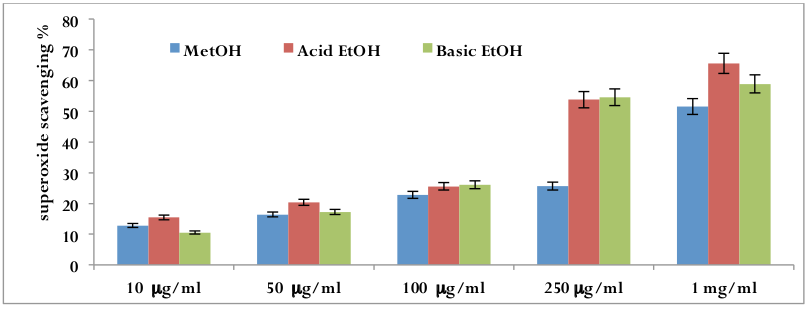
Photochemical reduction of flavins generates O 2 which reduces NBT, resulting in the formation of blue formazan Beauchamp and Fridovich, 1971. Three types of date extracts inhibited the formation of the blue formazan and % inhibition is proportional to the concentration [r2 =0.809 (p<0.01) for methanolic extract; r2 =0.908 (p<0.01) for acidic ethanolic extract; r2 =0.928 (p<0.01) for basic ethanolic extract]. IC50 values were 28.88 μμg/ml for methanolic extract, 19.00 μμg/ml for acidic ethanolic extract and 19.15 for acidic ethanolic extract. The superoxide ion scavenging activities of all these three extracts are: acidic ethanolic extract> basic ethanolic extract> methanolic extracts ( Figure 5 ).
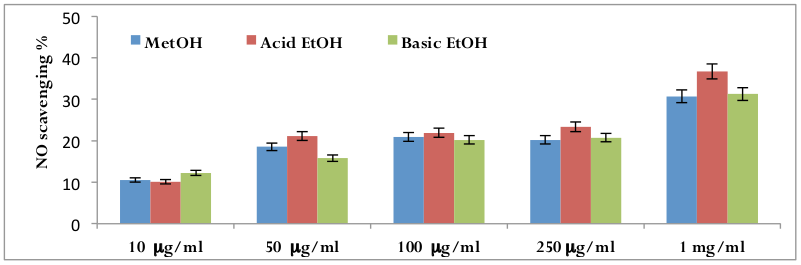
Nitric oxide (NO) scavenging activity
The calculated IC50 values of all three different date extracts suggested that methanolic extract is the most potent extract to scavenge the NO molecules followed by acidic ethanolic extracts and basic ethanolic extracts. But surprisingly there is little or no significant variation of NO radical scavenging properties of all three dateextracts ( Figure 6 ).
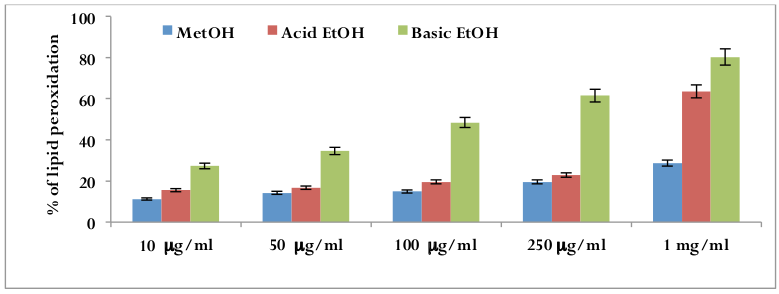
Lipid peroxidation inhibition assay
Egg yolk lipids undergo rapid nonenzymatic peroxidation when incubated in the presence of ferrous sulphate. Lipid peroxides are likely involved in numerous pathological events, including inflammation, metabolic disorders and cellular aging (Ames et al., 1993; Wiseman et al., 1996). Dateextracts inhibited lipid peroxidation in a concentrationdependent manner [r2=0.870(p<0.01) for methanolic extract; r2 =0.635 (p< 0.01) for acidic ethanolic extract; r2 =0.9596 (p<0.01) for basic ethanolic extract]. IC50 values for the inhibition of lipid peroxidation were 60.69 μμg/ml for methanolic extract, 24.75 μμg/ml for acidic ethanolic extract and 18.03 μμg/ml for basic ethanolic extract. The results suggested that consumption of date may afford a cytoprotective effect by lowering the lipid peroxidation level ( Figure 7 ).
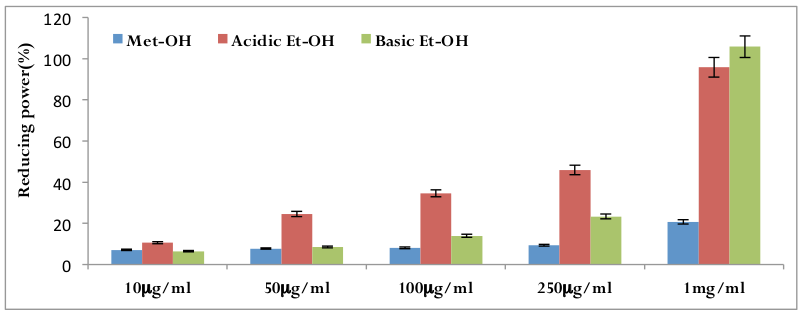
Determination of reducing power
The reducing power of different date extract samples using the potassium ferricyanide reduction method was evaluated. It has been investigated from the Fe 3+ –Fe 2+ transformation in the presence of three different extract samples. Highest activity was found in basic ethanolic extracts, followed by acidic ethanolic extracts of date and the lowest activity was found in methanolic extract of date ( Figure 8 ).
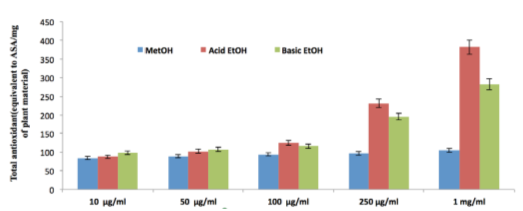
Total antioxidant capacity (equivalent to ASA/mg of plant material)
Total antioxidant capacity of date extract is expressed as the number of equivalents of ascorbic acid. The assay is based on the reduction of Mo(VI) to Mo(V) by the extract and subsequent formation of a green phosphate/Mo(V) complex at acid pH. The phosphomolybdenum method is quantitative since the antioxidant activity is expressed as the number of equivalents of ascorbic acid Prieto et al., 1999. Acidic ethanolic extract had a higher capacity than the other two varieties. The results from various free radicalscavenging system revealed that the three date extract samples collected by three different methods had significant antioxidant activity. The extracts were found to have different levels of antioxidant activity in the systems tested. The antioxidant activities of the three varieties were in the order acidic ethanolic extract>basic ethanolic extract>methanolic extract (Table 7) ( Figure 9 }).
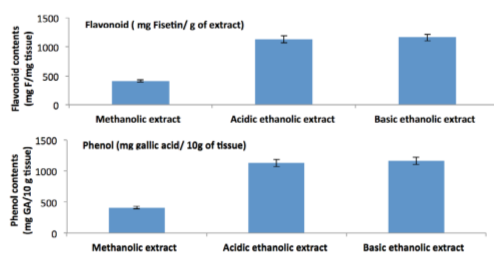
Total Flavonoid and phenol content.
Total flavonoid content expressed as Fisetin equivalents was 15.44±0.872 mg/g plant material in methanolic extract, 60.96±0.996 mg/g plant material in acid-‐ic ethanolic extract and 26.07±1.26 mg/g plant material in basic ethanolic extract respectively. Total phenolic concentration showed close correlation with the antioxidant activity, being highest in basic ethanolic extract and lowest in methanolic extract of date. Plant phenolics present in fruit and vegetables have received considerable attention because of their potential antioxidant activity (LópezVélez et al., 2003). Natural polyphenols have chainbreaking antioxidant activities and are believed to prevent many degenerative diseases, including cancer and atherosclerosis Roginsky, 2003, Table 8) ( Figure 10A,B ) Figure 10Flavonoid and Phenol contents in three date palm extracts(Though the basic ethanolic extract contain maximum phenolic contents than other two tested extracts, but the bioactive flavonoid content is highest in acidic ethanolic extracts. Methanolic date palm extract is very poor in containing phenol and flavonoids)/ Total flavonoid content of the date palm extracts was determined using aluminium chloride (AlCl 3 ) where as the total Phenolic content was determined using the Folin-Ciocalteu reagent. (n=5) results were expressed as mg of Flavonoid contents in fisetin/g of extracts and mg of gallic acid/ 10 mg of the extract, taking ±SD. Total Flavonoid and Phenol contents show a direct corelation with the total anti oxidant capacity of the tested compounds. Acidic ethanolic extract is rich in both Phenolic and Flavonoid contents, and it can be correlated to it’s radical scavenging activities and total antioxidant properties as the data has shown, than the other two tested extracts of date palm.)
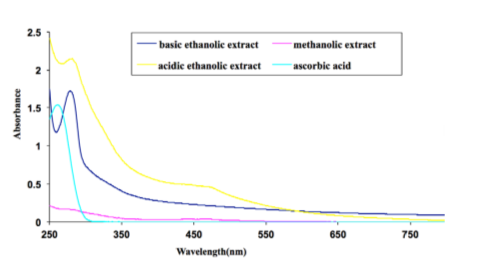
UV based spectrophotometric Analysis:
When different extracts of date was analysed in a UVVIS spectrophotometer to assess their solubility in different solvents to determine actual active compounds remain in that solution, we got a striking result where date extracts in basic ethanolic solvents yielded best solubility in comparison to their acidic ethanolic solvent, whereas the methanolic extracts failed to show any such significant peak, indicating very low or no solubility of the date extract into it which states that ethanolic solvent is the best to exhibit the potent anti inflammatory biological activities of the date which correlates with the cellular analysis further ( Figure 11 ).
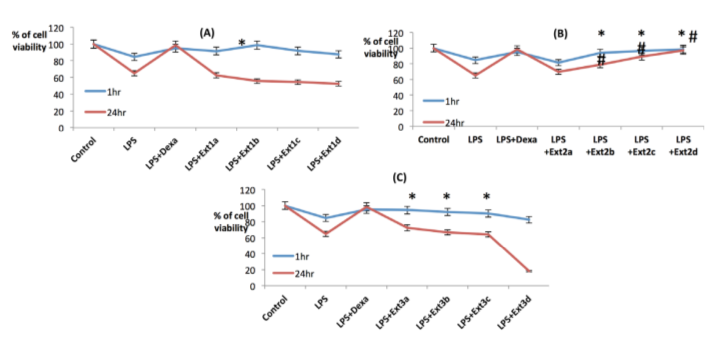
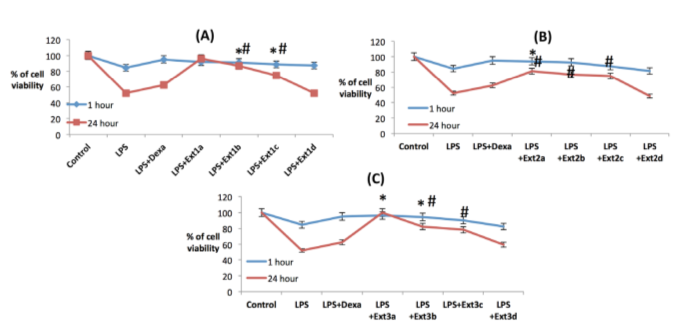
Cell viability of RAW macrophages:
In order to ascertain the antiinflammatory activities of the date extracts, collected by three distinct methodologies, LPS induced preclinical in vitro models, using both RAW macrophage like murine cell lines and HEK, human embryonic kidney cell lines were used. Cells were seeded into the 96 well plate and incubated for 12 hours, then 1 μμg/ml LPS was added into it following the various doses of the extracts and incubated for one hour and twenty four hour time periods respectively. In both the models using RAW macrophages, three extracts possess their anti inflammatory activities by regulating the cell viability in a dose dependent manner where methanolic extracts are not potent to maintain cell viability in 24 hour inflammatory models in comparison to the other two extracts where acidic ethanolic extracts retain the cell viability significantly in its lower doses, i.e. 100 μμg/ml, 50 μμg/ml, 10 μμg/ml and 1 μμg/ml respectively in both one hour and 24 hour models ( Figure 12a , Figure 12b ), but the basic ethanolic extracts maintain the cell viability only in higher doses, i.e. 250 μμg/ml, 100 μμg/ml and 50 μμg/ml, where in lower doses (1 μμg/ml) it acts as proinflammatory causing the cell death ( Figure 12c ). So all these extract may act as either antiinflammatory or pro inflammatory depending on their doses and nature of extraction.
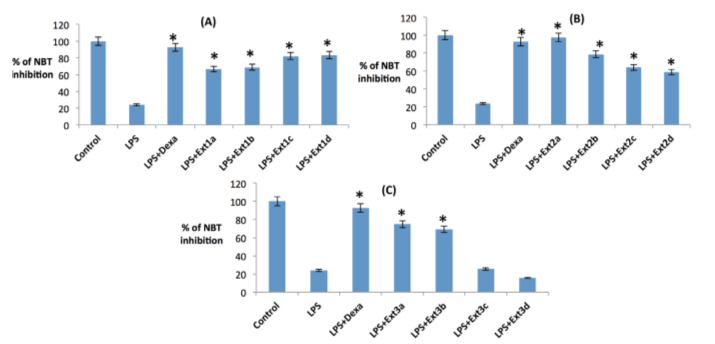
Assay of hydroxyl radical (OH) scavenging activity
As the in vitro study suggested that all three date crude extracts have potent hydroxyl radical and superoxide ion scavenging activities, the ex vivo result, using RAW macrophages, also advocated the same where acidic ethanolic extract has been shown to have strongest hydroxyl radical scavenging power, >50% inhibition ( Figure 13b ), followed by basic ethanolic ( Figure 13c ) and methanolic extracts ( Figure 13a ). This mayhave been accomplishedby modulating the mitochondria associated NADPH oxidase activity.
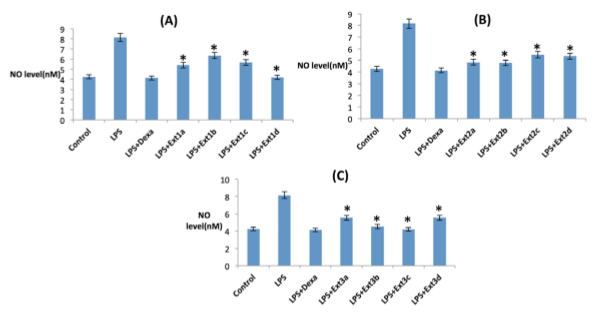
NO estimation
None of the tested compounds induced changes in NO basal levels, when incubated without LPS (data not shown). To evaluate the nitrosative stress, Griess reagent was used. After24 h LPS treatment, cellular NO production increases nearly two folds (Figure 14a) in comparison to the untreated control samples, whereas the three date extracts ameliorate the LPS induced NO production to the basal level like untreated group in a dose dependent manner which strongly showing there NO scavenging capabilities which is already been established by the in vitro assays. Here dexamethasone has been taken as positive control ( Figure 14a,b,c ).
Cell viability of HEK 293 cells
To investigate the cytotoxicity of the three date extracts MTT assay was performed as MTT is reduced bymitochondrial dehydrogenase to form formazan, an insoluble purple compound and one can measure the cytotoxicity in terms of the intensity of the purple compound. Whereas on the other hand, dead cells do not form any purple formazan because the enzyme is degraded and lack regular function. Among the three extracts of date, acidic ethanolic extracts ameliorated the LPS induced cell death to retain the cell viability in both one hour and twenty four hour inflammatory models ( Figure 15b ) followed by basic ethanolic ( Figure 15c ) and methanolic extracts ( Figure 15a ) in a dose dependent manner. In lower doses all these three extracts fail to inhibit LPS induced cell death where as they are active only in the higher concentrations, i.e. in 250 μμg/ml and 100 μμg/ml doses which is similar to the LPS induced inflammatory models applied on murine macrophage cell line. This data suggests that acidic ethanolic extracts contain potent antiinflammatory compound(s) which is/are able to inhibit the LPS induced cytotoxicity in a dose dependent manner. Dexamethasone was used as positive control in the entire study.
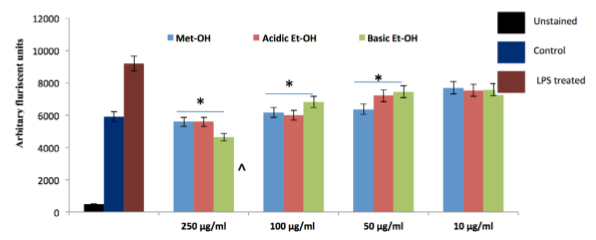
ROS measurement
The evidence presented above suggests that date extracts mediates its effects through the antioxidant pathway. We used a DCFDA probe to examine whether this mechanism can scavenge ROS inside the RAW macrophage cells. Cells were treated with E.coli LPS and then various concentrations of date extracts were added into it therapeutically and incubated for 6 h, and analyzed by florescent spectrophotometer. Date extracts ameliorate LPS induced ROS levels significantly over the control in a dose dependent manner. This effect was observed best in acidic ethanolic extracts where it inhibited the ROS generation in higher doses, i.e. in 250 μμg/ml and 100 μμg/ml and increased ROS levels slightly thereafter. ROS levels decreased significantly in methanolic and basic ethanolic extracts also but increased significantly in its lower doses compared to control cells ( Figure 16 ).

Measurement of mitochondrial membrane potential (MMP) After the assessment of total cellular antioxidant property, in the next phase we targeted mitochondria, the most important site inside the cell for ROS generation. Here we have found that all three extracts of date is capable of reducing LPS induced oxidative stress in murine macrophage like cells significantly, in comparison to the control group in a dose dependent manner, which advocates that the date extracts ameliorate the LPS induced oxidative stress by targeting the mitochondria in a dose dependent manner and thus maintain the cell viability ( Figure 17 ).
TLC analyses
Samples Sample Samples
Sample A: The extract A showed several peaks as shown in the Figure 18 .However only two peaks show appreciably high absorbance when observed with 254 nm wavelength. The peak at 23.5 min is the major peak while that at 34.0 min is a minor peak. The peak at 23.5 min appears about 20 fold attenuated when observed with 370 nm wavelength while the peak at 34.0 min is not observed at this wavelength. Sample B: Extract B shows four closely spaced peaks between 6.0 – 9.0 min appearing at 6.7, 7.3, 7.8 and 8.4 min with major peak being at 7.3 min when observed with 254 nm wavelength. These peaks were also observed with 370 nm wavelength but with about 4 fold attenuation in peak absorbance values. Sample C: The extract C showed roughly three distinct peaks. They were at 5.15.2 min, 6.3 min and 7.9 min using 254 nm wavelength. The first two peaks are major peaks but are not well separated. The third peak at 7.9 min is a minor peak in this sample. There is another minor peak at about 6.8 min observed with 254 nm wavelength which may be a spurious peak as it lies on the shoulder of the 6.3min peak and didn’t appear in repeat scan. The absorbance maxima of these peak positions remained similar when seen in 370nm wavelength although their intensities were attenuated by about 10 fold. Comparing the absolute absorbance values of peak maxima of sample C with others samples (A &B), it indicates that sample C contains much higher concentration of compound(s) as similar values of samples (100200 μl) were injected for HPLC analysis in each case.

HPLC analyses
Figure 19 Figure 19 HPLC profile of basic ethanolic extract
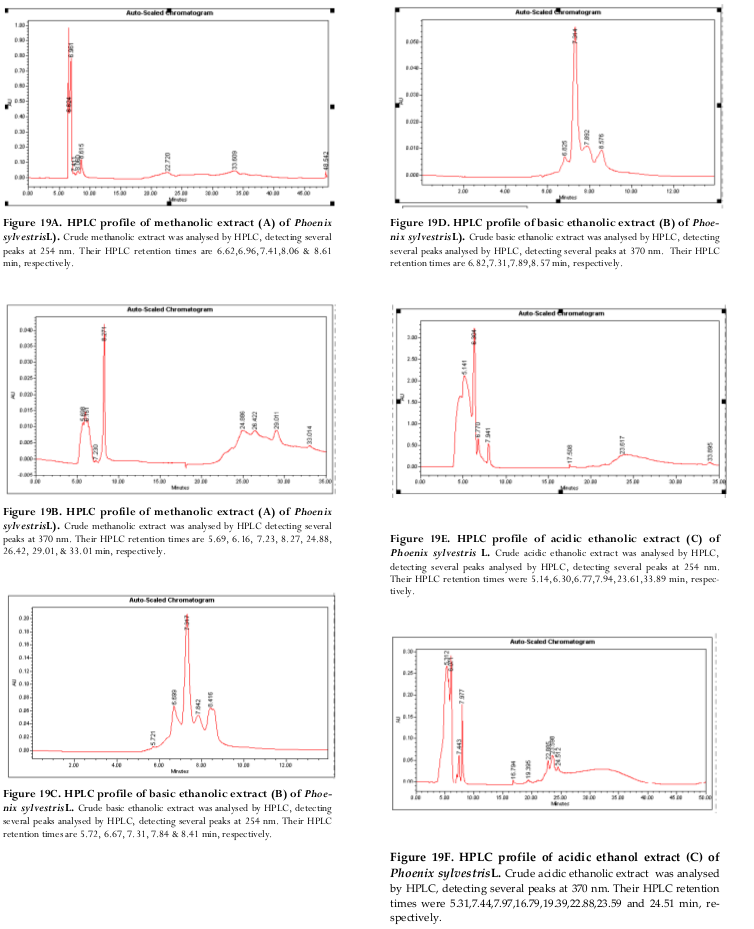
Discussion
In the present study to evaluate antiinflammatory potential of the extracts of date collected by three different extraction procedures ( Figure 1 ) where one or more fractions of the mesocarp tissue is likely to be included and the rest excluded, various inflammatory markers were used against a powerful and well char acterized proinflammatory stimulusLPS, a major component of the outer membrane of Gramnegative bacteria. This molecule shows potent pro inflammato ry action on various cell types, including macrophages, endothelial cells, and fibroblasts Allaith, 2008 induced inflammation in RAW 264.7 and HEK 291 cell lines by interacting with TLR4, a transmembrane receptor on macrophages, which recognizes molecules derived from gram negative bacteria activating the downstream NFκB (nuclear factor kappa) signaling
pathway leading to increased production of cytokines, and/or ROS/RNS generation following the activation of mitogenactivated protein kinases (MAPK) and NFκB mediated signaling pathway which ultimately drive the tissue degeneration.
Fruits constitute an important part of a balanced diet as they are natural sources of food nutrient needed by human and animals, being rich in proteins, carbohydrates, minerals and dietary fibers. Phoenix dactylifera, belongs to tree palm family Arecaceae, consumed throughout the world and is an important part of the diet in the Middle East Barh and Mazumdar, 2008. Phytochemical screening has revealed dried date pollens to contain sterols and other supplements such as vitamins and cofactors Ishurd et al., 2003 and lack volatile substances Hussein et al., 1998. Phytochemi cal analyses of fruit flesh and seed of date revealed the presence of flavonoids (luteolin, methyl luteolin, quer
cetin, and methyl quercetin), flavanols (catechin, epicatechin) (Alshahib and Marshall, 2003; Bennett et al., 1966)Heftmann et al., 1965, free phenolic acids (protocatechuic acid, vanillic acid, syringic acid, and ferulic acid) (Mahran et al., 1976; Tabeta et al., 2002), tan nins, glycosides, cardiac glycosides and steroids (cholesterol, stigmasterol, campesterol and αsitosterol), essential amino acids Biglari et al., 2008Hong et al., 2006Ziouti et al., 1996; and nine bound phenolic acids (gallic acid, protocatechuic acid, phydroxybenzoic acid, vanillic acid, caffeic acid, syringic acid, pcoumaric acid, ferulic acid, and ocoumaric acid) Hong et al., 2006Hussein et al., 1998 respectively. In the past few years, many pharmacological studies have been conducted on Phoenix dactylifera (Abdulla; AlQarawi et al., 2003; Ishurd and Kennedy, 2005). Various in vitro and in vivo antioxidant assays have been carried out on various extracts of different parts of Phoenix dactylifera Vayalil, 2002. Oral administra tion of the methanolic and aqueous extracts of edible portion of Phoenix dactylifera fruits suppressed swelling in the foot, while the methanolic extract of date seeds showed significant reduction in adjuvant arthritis in rats Bastway Ahmed et al., 2010 and on gentamicin induced nephrotoxicity in rats Javanmardi, 2003.
In the present study, datefruit was extracted using various organic solvents, and their antioxidative and antiinflammatory activities were evaluated. Phenolic compounds as well as Flavonoids, which are a group of poly phenolic compounds possessing free radical scavenging properties, potential inhibitory actions of hydrolytic and oxidative enzymes as well as anti inflammatory action. Obtained data have clearly demonstrated that date extracts, depending on their mode of collectionpossess potent anti oxidative properties indicated by significant inhibition of lipid peroxidation ( Figure 7 ), superoxide radical scavenging, hydroxyl radical scavenging, DPPH radical scavenging and nitric oxide (NO) scavenging activities. The calculated IC50 values of dateindicate strong anti oxidative properties of these crude extracts.
Data obtained from studies using these date extracts, collected by three different methods indicate potentanti oxidative properties by showing significant inhibition of lipid peroxidation, superoxide radical scavenging, hydroxyl radical scavenging, DPPH radical scavenging and nitric oxide (NO) scavenging activities. The calculated IC50 valuesof these three extracts of datehaving revealedstrong antioxidative properties ( Figure 9 , Table 7), their anti inflammatory potencies warranted confirmation through additional studies on cells. Both human and murine cell lines were assessed. Oxidative stress being the main driving force in anyinflammatory cascade, studies weredesigned to evaluate theirameliorative effect during and postoxidative stress induced inflammation.
As we have identified the anti oxidative capacity ( Figure 9 ) of dietary date extracts, collected by three different extraction procedures, i.e. methanolic, basic ethanolic and acidic ethanolic methods, ( Figure 1 ) as an interesting lead that can stabilize the intracellular reactive oxygen and nitrogen intermediates in both murine and human cell lines in vitro at non cytotoxic concentrations, we were therefore interested to evaluate the in vivo antiinflammatory activities of these compounds in preclinical models, but before this cytotoxic and anti inflammatory properties of these three extracts have been evaluated thoroughly, targeting the mitochondrial reactive oxygen and nitrogen intermediates ( Figure 17 ).
The acidic ethanolic extracts of date extract amelio rates the LPS induced oxidative stress by inhibiting the NO production ( Figure 6 ) and subsequently protect ing the mitochondrial respiratory chain complex function, thus maintaining the cellular homeostasis or cell viability per se. As we have mentioned earlier that oxidative stress and mitochondrial dysfunction are key features for any inflammatory cascade and here the date extracts are capable of inhibiting this, thus, these crude extracts must have potent anti inflammatory power. Another important point to be noted here is that the acidic ethanolic extract of the date has shown a more potent antiinflammatory properties in murine RAW macrophages ( Figure 12b ) in comparison to other two extracts ( Figure 12a Figure 12c ), whereas in case of LPS induced human HEK cells, the methanolic extract ( Figure 15a ) is better at inhibiting the LPS induced oxidative stress and in maintaining the cellular homeostasis, than either acidic or basic etha nolic extracts. From studies done by various researchers as stated and referred to in the preceding paragraphs, we now know that plants have various phenolic compounds and flavonoids, which are basically secondary metabolites of the pentose phosphate, shikimate, and phenylpropanoid pathways, exhibit strong anti oxidative redox properties allowing them to act as reducing agents, hydrogen donors, and singlet oxygen quenchers. Also, many of them have been shown to exhibit the antagonism for the classical NFκB signaling pathways and for this reason in past few years there has been an upsurge of interest in the therapeutic potentials of medicinal plants as antioxidants in reducing such free radical induced tissue injury.
Although several phytochemicals have been shown to possess pharmacological properties of potential interest antiinflammatory properties and/or therapy, their activity in the in vitro and in vivo preclinical model organisms is still not well understood due to variations of their extraction and collection procedures, heterogeneous solubility in different solvents and most importantly, route of administration of these compounds in a dose dependent manner in different disease models. The mechanism by which various date extracts mediates its antioxidant effects remains unclear.
Mitochondria are the major source for ROS generation in the cell. Evidence from our studies suggests the role of mitochondria in date induced ROS scavenging pathways ( Figure 17 ). It is possible that date extracts in hibit the mitochondrial enzymes that lead to production of ROS and thus maintain the oxidative stress induced cell damage ( Figure 16 ). The inhibition of ROS by date extracts could occur through its interaction with thioredoxin reductase thus changing its activity to NADPH oxidase, which could then lead to the scavenging of ROS. It is not clear yet which structural group or compound of date extracts is responsible for inhibiting the ROS production. Present extracts of date also has shown a high level of phenol ( Figure 10b ) and flavonoid ( Figure 10a ) content and this may be the reason behind their antiinflammatory activities.
We can conclude that the inhibitory effect of date extracts on LPS induced inflammation in RAW macrophages is via the downstreaming of the classical TLR4 mediated signaling cascades scavenging the mitochondrial ROS production ( Figure 17 ). Until phytochemical screening of these extracts is carried out, this will be very early to conclude. However, it is true that the compounds present in these extracts can inhibit the endotoxin induced oxidative stress ( Figure 16 ) in both murine and human cell lines in a dose dependent manner. The phytochemical screening and the structure activity relationship of these extracts are under investigation now along with the detailed antiinflammatorysignaling pathways. But the question is, what compound (s) are exactly present in these extracts that are truly exhibiting the antiinflammatory properties and the signaling pathways that they follow, is still not clear yet. For this HPLC and massspectrometry of the samples are being carried out and from database analysis individual eluents shall be used in antiinflammatory and antioxidative assays to assess their oxiflammatory potential. This is ongoing work in the lab.
Acting as second messengers, transient free radicals, synthesized during regular metabolismoften trigger further downstream sequence of activation networks. But produced in unregulated catabolic cycles, they demonstrate deleterious compounds such as toxins and wastes which further contribute to the disease etiology. Studies on intermediate free radicals, especially reactive oxygen species (ROS) and their action on cellular physiology, have shown that they play a pivotal role in causing secondary tissue degeneration in various inflammatory diseases, such as rheumatoid arthritis Filippin et al., 2008, multiple sclerosis Offen et al., 2004, thyroiditis Burek and Rose, 2008, and type 1 diabetes Chen et al., 2008. ROS are known to perform essential roles in immune response to pathogens, including bacterial killing via induction of superoxide anion during respiratory burst in activated macrophages and neutrophils Kanayama and Miyamoto, 2007Lambeth, 2004. Further studies on patients with chronic granulomatous disease (CGD) or genetically engineered mice lacking components of the NADPH oxidase enzyme (NOX) Morgenstern, 1997Shiloh et al., 1999 provide corroborative data.
Reactive oxygen species (ROS) oxidativelymodify DNA, proteins, lipids, and small intracellular molecules. Lipids, for example, pulmonary surfactants, react with ROS to produce lipid peroxides that perpetrate increased membrane permeability and inactivation of surfactants Yang et al., 2006. Further, ROS react with cellular proteins and inhibit protein synthesis by disabling proteins involved in translation and translocation. This effectively impair cellular metabolism Tuder et al., 2003. ROS also damage nucleic acids by modifying purine and pyrimidine bases and by causing DNA strand breakage Dizdaroglu et al., 2002. Overproduction of ROS and oxidative stress has been found to be critical in pathophysiology of other complex syndromeslike cancer, asthma, cystic fibrosis, ischemiareperfusion injury, druginduced toxicity, and aging Jezierska-Drutel et al., 2013. Natural products have yielded as many as 70% of the drugs used today for inflammation and degeneration Newman et al., 2002. Consumption of large portions of fruits and vegetables have been recommended for reduction of risk of cardiovascular diseases, autoimmune diseases, cancer, and various other chronic illnesses Neuhouser, 2004 although little is known about the actual active compounds or moieties which are beneficial. It is an important project to identify and validate direct action and participation of such active ingredients in modulating disease.
Figures 18-20 show data on proteomic analyses by TLC, HPLC and MS. On preliminary investigation TLC data reveals that extract A may not contain important compounds although antiinflammatory activity was seen in the other two extracts. Clear peaks were not obtained in scan data of the other two which necessitates further purification steps and MS analyses of the individual peaks and NMR. This work is ongoing and is beyond the scope of this work.
Conclusion
In this study, three different types of date extracts, methanolic, acidic ethanolic and basic ethanolic produce effects on reactiveoxygen species where scavenging of hydroxyl radicals, superoxide radicals, nitric oxide scavenging activity, and inhibition of lipid peroxidation were found to occur in a dose dependent manner. Moreover, they have a significant inhibition of antiinflammatory activities. Invitro experiments prove that the effects of the extracts islikely mediated through inhibition of mitochondria derived (ROS) scavenging pathways. Thisis an important finding as it shows not only the extracts’ pathway of action but also provesthe direct involvement of a subcellular signaling network that can be interfered with, using anatural products without the possibility of drastic side effects. Ongoing proteomic analysis has already revealed scan data using various sensitive chromatographic techniques of certain specific peaks. Further analyses are needed to conclusively identify the peaks to reveal identity of one or more compounds for detailed characterization and development into novel drug entities.
Abbreviations
Human embryonic kidney cell line (HEK); murine RAW macrophages; lipopolysaccharide (LPS); reactive oxygen species (ROS); nitric oxide (NO); 1,1-Diphenyl-2-picrylhydrazyl radical (DPPH); dihydro rhodamine (DHR); super oxide dismutase (SOD).
References
-
Y. Al-Taher
Abdulla.
Possible anti-diarrhoeal effect of the date palm (Phoenix Dactylifera L) spathe aqueous extract in rats. Scientific Journal of King Faisal University (Basic and Applied Sciences).
2008;
9
:
1429-1435
.
-
A.A.
Al-Qarawi,
B.H.
Ali,
S.A.
Al-Mougy,
H.M.
Mousa.
Gastrointestinal transit in mice treated with various extracts of date (Phoenix dactylifera L.). Food and Chemical Toxicology.
2003;
41
:
37-39
.
-
W.
Al-shahib,
R.J.
Marshall.
The fruit of the date palm: its possible use as the best food for the future?. Int J Food Sci Nutr.
2003;
54
:
247-259
.
-
A.A.A.
Allaith.
Antioxidant activity of Bahraini date palm (Phoenix dactylifera L.) fruit of various cultivars.. International Journal of Food Science & Technology.
2008;
43
:
1033-1040
.
-
R.
Amann,
R.
Schuligoi,
I.
Lanz,
J.
Donnerer.
Histamine-induced edema in the rat paw — effect of capsaicin denervation and a CGRP receptor antagonist. European Journal of Pharmacology.
1995;
279
:
227-231
.
-
D.
Barh,
B.
Mazumdar.
Comparative nutritive values of palm saps before and after their partial fermentation and effective use of wild date (Phoenix sylvestris Roxb.) sap in treatment of anemia.. Research Journal of Medicine and Medical Sciences.
2008;
3
:
173-176
.
-
M.
Bastway Ahmed,
N.
Hasona,
A.
Selemain.
Protective effects of extract from dates (Phoenix dactylifera L.) and ascorbic acid on thioacetamide-induced hepatotoxicity in rats. Iranian Journal of Pharmaceutical Research.
2010;
:
193-201
.
-
C.
Beauchamp,
I.
Fridovich.
Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analytical Biochemistry.
1971;
44
:
276-287
.
-
R.D.
Bennett,
S.-t.
Ko,
E.
Heftmann.
Isolation of estrone and cholesterol from the date palm, Phoenix dactylifera L. Phytochemistry.
1966;
5
:
231-235
.
-
F.
Biglari,
A.F.M.
AlKarkhi,
A.M.
Easa.
Antioxidant activity and phenolic content of various date palm (Phoenix dactylifera) fruits from Iran. Food Chemistry.
2008;
107
:
1636-1641
.
-
A.
Braca,
N.
De Tommasi,
L.
Di Bari,
C.
Pizza,
M.
Politi,
I.
Morelli.
Antioxidant Principles from Bauhinia t arapotensis. J Nat Prod.
2001;
64
:
892-895
.
-
C.L.
Burek,
N.R.
Rose.
Autoimmune thyroiditis and ROS. Autoimmun Rev.
2008;
7
:
530-537
.
-
J.A.
Chatpalliwar VA,
MM
Wanjari,
RR
Chakraborty,
VT
Kharkar.
Antiinflammatory activity of Martynia diandra GLOX. Indian Drugs.
2002;
39(10)
.
-
J.
Chen,
A.M.
Gusdon,
T.C.
Thayer,
C.E.
Mathews.
Role of Increased ROS Dissipation in Prevention of T1D. Annals of the New York Academy of Sciences.
2008;
1150
:
157-166
.
-
S.-K.
Chung,
T.
Osawa,
S.
Kawakishi.
Hydroxyl Radical-scavenging Effects of Spices and Scavengers from Brown Mustard (Brassica nigra ). Bioscience Biotechnology and Biochemistry.
1997;
61
:
118-123
.
-
R.a.
Cortran.
Acute and Chronic Inflammation.. In Robbins and Cotran Pathologic Basis of Disease (Elsevier BV).
2010;
:
43-77
.
-
P.
Crunkhorn,
S.C.R.
Meacock.
Mediators of the inflammation induced in the rat paw by carrageenin. British Journal of Pharmacology.
1971;
42
:
392-402
.
-
M.
Dizdaroglu,
P.
Jaruga,
M.
Birincioglu,
H.
Rodriguez.
Free radical-induced damage to DNA: mechanisms and measurement 1, 2. Free Radical Biology and Medicine.
2002;
32
:
1102-1115
.
-
L.I.
Filippin,
R.
Vercelino,
N.P.
Marroni,
R.M.
Xavier.
Redox signalling and the inflammatory response in rheumatoid arthritis. Clinical & Experimental Immunology.
2008;
152
:
415-422
.
-
J.
Gutteridge.
Ferrous-salt-promoted damage to deoxyribose and benzoate. The increased effectiveness of hydroxyl-radical scavengers in the presence of EDTA. Biochem J.
1987;
243
:
709-714
.
-
J.M.C.
Gutteridge.
Adriamycin-iron catalysed phospholipid peroxidation: a reaction not involving reduced adriamycin or hydroxyl radicals. Biochemical Pharmacology.
1983;
32
:
1949-1952
.
-
E.
Heftmann,
S.-T.
Ko,
R.D.
Bennett.
Identification of estrone in date seeds by thin-layer chromatography. Naturwissenschaften.
1965;
52
:
431-432
.
-
Y.J.
Hong,
F.A.
Tomas-Barberan,
A.A.
Kader,
A.E.
Mitchell.
The Flavonoid Glycosides and Procyanidin Composition of Deglet Noor Dates ( Phoenix dactylifera ). Journal of Agricultural and Food Chemistry.
2006;
54
:
2405-2411
.
-
M.M.-D.
Hussein,
W.A.
Helmy,
H.
Salem.
Biological activities of some galactomannans and their sulfated derivatives. Phytochemistry.
1998;
48
:
479-484
.
-
O.
Ishurd,
Y.
Ali,
W.
Wei,
F.
Bashir,
A.
Ali,
A.
Ashour,
Y.
Pan.
An alkali-soluble heteroxylan from seeds of Phoenix dactylifera L. Carbohydrate Research.
2003;
338
:
1609-1612
.
-
O.
Ishurd,
J.F.
Kennedy.
The anti-cancer activity of polysaccharide prepared from Libyan dates (Phoenix dactylifera L.). Carbohydrate Polymers.
2005;
59
:
531-535
.
-
J.
Javanmardi.
Antioxidant activity and total phenolic content of Iranian Ocimum accessions. Food Chemistry.
2003;
83
:
547-550
.
-
A.
Jezierska-Drutel,
S.A.
Rosenzweig,
C.A.
Neumann.
Role of oxidative stress and the microenvironment in breast cancer development and progression. Advances in cancer research.
2013;
119
:
107
.
-
A.
Kanayama,
Y.
Miyamoto.
Apoptosis triggered by phagocytosis-related oxidative stress through FLIPS down-regulation and JNK activation. Journal of Leukocyte Biology.
2007;
82
:
1344-1352
.
-
J.D.
Lambeth.
NOX enzymes and the biology of reactive oxygen. Nature Reviews Immunology.
2004;
4
:
181-189
.
-
M.
López-Vélez,
F.
Martínez-Martínez,
C.D.
Valle-Ribes.
The Study of Phenolic Compounds as Natural Antioxidants in Wine. Critical Reviews in Food Science and Nutrition.
2003;
43
:
233-244
.
-
G.
Mahran,
S.
Abdel-Wahab,
A.
Attia.
A PHYTOCHEMICAL STUDY OF DATE PALM POLLEN. Planta Med.
1976;
29
:
171-175
.
-
C.A.
Martinez,
M.E.
Loureiro,
M.A.
Oliva,
M.
Maestri.
Differential responses of superoxide dismutase in freezing resistant Solanum curtilobum and freezing sensitive Solanum tuberosum subjected to oxidative and water stress. Plant Science.
2001;
160
:
505-515
.
-
A.A.
Miles,
E.M.
Miles.
Vascular reactions to histamine, histamine-liberator and leukotaxine in the skin of guinea-pigs. The Journal of Physiology.
1952;
118
:
228-257
.
-
H.
Mohan.
Chapter-06 Inflammation and Healing.. In Textbook of Pathology (Jaypee Brothers Medical Publishing).
2010;
:
130-173
.
-
D.E.
Morgenstern.
Absence of Respiratory Burst in X-linked Chronic Granulomatous Disease Mice Leads to Abnormalities in Both Host Defense and Inflammatory Response to Aspergillus fumigatus. Journal of Experimental Medicine.
1997;
185
:
207-218
.
-
G.H.
Naik,
K.I.
Priyadarsini,
J.G.
Satav,
M.M.
Banavalikar,
D.P.
Sohoni,
M.K.
Biyani,
H.
Mohan.
Comparative antioxidant activity of individual herbal components used in Ayurvedic medicine. Phytochemistry.
2003;
63
:
97-104
.
-
M.L.
Neuhouser.
Dietary flavonoids and cancer risk: evidence from human population studies. Nutr Cancer.
2004;
50
:
1-7
.
-
D.J.
Newman,
G.M.
Cragg,
S.
Holbeck,
E.A.
Sausville.
Natural products and derivatives as leads to cell cycle pathway targets in cancer chemotherapy. Curr Cancer Drug Targets.
2002;
2
:
279-308
.
-
D.
Offen,
Y.
Gilgun-Sherki,
E.
Melamed.
The role of oxidative stress in the pathogenesis of multiple sclerosis: The need for effective antioxidant therapy. Journal of Neurology.
2004;
251
:
261-268
.
-
H.
Ohkawa,
N.
Ohishi,
K.
Yagi.
Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry.
1979;
95
:
351-358
.
-
P.
Prieto,
M.
Pineda,
M.
Aguilar.
Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E. Analytical Biochemistry.
1999;
269
:
337-341
.
-
V.
Roginsky.
Chain-breaking antioxidant activity of natural polyphenols as determined during the chain oxidation of methyl linoleate in Triton X-100 micelles. Archives of Biochemistry and Biophysics.
2003;
414
:
261-270
.
-
G.
Ruberto,
M.T.
Baratta,
S.G.
Deans,
H.J.D.
Dorman.
Antioxidant and Antimicrobial Activity of Foeniculum vulgare and Crithmum maritimum Essential Oils. Planta med.
2000;
66
:
687-693
.
-
N.
Salah,
N.J.
Miller,
G.
Paganga,
L.
Tijburg,
G.P.
Bolwell,
C.
Riceevans.
Polyphenolic Flavanols as Scavengers of Aqueous Phase Radicals and as Chain-Breaking Antioxidants. Archives of Biochemistry and Biophysics.
1995;
322
:
339-346
.
-
M.U.
Shiloh,
J.D.
MacMicking,
S.
Nicholson,
J.E.
Brause,
S.
Potter,
M.
Marino,
F.
Fang,
M.
Dinauer,
C.
Nathan.
Phenotype of Mice and Macrophages Deficient in Both Phagocyte Oxidase and Inducible Nitric Oxide Synthase. Immunity.
1999;
10
:
29-38
.
-
S.
Tabeta,
I.O.
Uehare,
M.
Zahid,
H.
Zhou,
P.
Yoshioka.
Yuanjiang. α-D-glucan structure. Carbohydrate Research.
2002;
337
:
1325-1328
.
-
R.M.
Tuder,
L.
Zhen,
C.Y.
Cho,
L.
Taraseviciene-Stewart,
Y.
Kasahara,
D.
Salvemini,
N.F.
Voelkel,
S.C.
Flores.
Oxidative stress and apoptosis interact and cause emphysema due to vascular endothelial growth factor receptor blockade. Am J Respir Cell Mol Biol.
2003;
29
:
88-97
.
-
P.K.
Vayalil.
Antioxidant and Antimutagenic Properties of Aqueous Extract of Date Fruit ( Phoenix dactylifera L. Arecaceae).. Journal of Agricultural and Food Chemistry.
2002;
50
:
610-617
.
-
R.
Vinegar,
W.
Schreiber,
R.
Hugo.
Biphasic development of carrageenin edema in rats. J Pharmacol Exp Ther.
1969;
166
:
96-103
.
-
B.A.
Whittle.
THE USE OF CHANGES IN CAPILLARY PERMEABILITY IN MICE TO DISTINGUISH BETWEEN NARCOTIC AND NONNARCOTIC ANALGESICS. British Journal of Pharmacology and Chemotherapy.
1964;
22
:
246-253
.
-
W.
Wijesinghe,
M.
Senevirathne,
M.-C.
Oh,
Y.-J.
Jeon.
Protective effect of methanol extract from citrus press cakes prepared by far-infrared radiation drying on H2O2-mediated oxidative damage in Vero cells. Nutrition research and practice.
2011;
5
:
389-395
.
-
C.A.
Winter,
E.A.
Risley,
G.W.
Nuss.
Carrageenin-Induced Edema in Hind Paw of the Rat as an Assay for Antiinflammatory Drugs. Experimental Biology and Medicine.
1962;
111
:
544-547
.
-
Q.
Yang,
Y.-S.
Kim,
Y.
Lin,
J.
Lewis,
L.
Neckers,
Z.-G.
Liu.
Tumour necrosis factor receptor 1 mediates endoplasmic reticulum stress-induced activation of the MAP kinase JNK. EMBO Rep.
2006
.
-
A.
Ziouti,
C.
El Modafar,
A.
Fleuriet,
S.
El Boustani,
J.J.
Macheix.
Phenolic compounds in date palm cultivars sensitive and resistant to Fusarium oxysporum. Biologia Plantarum.
1996;
38
:
451-457
.
Comments

Downloads
Article Details
Volume & Issue : Vol 2 No 06 (2015)
Page No.: 297-317
Published on: 2015-06-17
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 8548 times
- Download PDF downloaded - 2289 times
- View Article downloaded - 8 times

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}