

Copyrights: Mohammad Reza Khosravi, Mohammad Hossein Mohammadi, Parinaz Khadem, Sahar Lashkari, Hamideh Aghaeenezhad, Ahmad Gharehbaghian, Zaher Khazaei, Mehdi Allahbakhshian Farsani, 2018. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Background: Acute lymphoblastic leukemia (ALL) arises from an imbalanced proliferation and differentiation of lymphoid progenitors due to special chromosomal and epigenetic abnormalities affecting cell cycle regulation. The cyclin-dependent kinase inhibitor (CDKI) family has crucial functions in G1 progression and G1 to S entry regulation. Among CDKIs, P21 and P27 are able to exert remarkable effects on all CDKs. Hence, we investigated the expression levels of P21 and P27 in ALL patients to determine whether or not their expression had been altered.
Materials and Methods: In the present study, we evaluated P21 and P27 expression in bone marrow and peripheral blood samples of 52 newly diagnosed ALL patients (30 males, 22 females) and 13 healthy normal controls (5 males, 8 females) using quantitative real-time PCR. Data were analyzed via SPSS (version 16) software and P<0.05 was assigned as the statistical significance level.
Results: Our findings demonstrated lower expression levels of P21 and P27 in ALL patients compared with normal controls (8.33- and 1.69-fold change, respectively). P21 and P27 expression was significantly different between T-cell acute lymphoblastic leukemia (T-ALL) and B-cell acute lymphoblastic leukemia (B-ALL) patients (P= 0.03).
Conclusion: Since P21 and P27 are able to influence the activity of both cyclins and CDKs, it is postulated that decreased expression of these genes reduces P21- and P27- mediated suppressive effects on cyclins and CDKs. Therefore, these events facilitate the activation of cyclins and CDKs which may result in cancer progression in ALL patients.
Background
Acute lymphoblastic leukemia (ALL) originates from blocked differentiation of lymphoid progenitors and affects both children and adults 1. Recent studies have tried to reveal the precise events contributing to the pathogenesis of ALL in order to enhance the effectiveness of therapeutic strategies in high-risk adult and childhood patients 2. Generally, ALL arises due to genetic lesions in genes involved in cell cycle regulation, apoptosis, and survival 3. Cell cycle regulators consist of families of cyclin-dependent kinase (CDK) inhibitors (CDKIs), including INK4 (p15, p16, p18, and p19) and CIP/KIP (P21CIP, P27KIP1, and p57KIP2) 4.
P21Cip1 and P27Kip1 are crucial CDK2 inhibitors which prevent G1 to S entry. The regulation of CDKI expression is considered as a common mechanism for cell integrity. In this regard, P21 or P27 over-expression acts as a vital component in tumor growth suppression. Furthermore, P21 and P27 are implicated in senescence and terminal differentiation of various types of cells, including hematopoietic cells. Several studies have demonstrated that P21 knockout mice are affected by different tumors, such as sarcomas and lymphomas. However, P21 gene mutations are infrequently detected in cancer cells. Moreover, decreased expression of P27 has been reported in a variety of cancers and contributes to tumor development 5. In primary T-ALL cells, P27 downregulation was associated with cell cycle progression and apoptosis inhibition 6.
Interestingly, recent studies have indicated a correlation between DNA methylation and decreased expression of some KIP family genes, such as P21, in ALL patients. Nevertheless, promoter methylation pattern of KIP family genes indicated inconsistent results. Traditionally, DNA damage induces cell cycle arrest via P21 and P27 activation. Although P21 and P27 have been considered as tumor suppressors in some studies, they have emerged as potential oncogenes in other studies as well. More studies have indicated that P21 and P27 could have additional activity beyond their function as CDK inhibitors. In this regard, Cip/Kip proteins might play a crucial role in the regulation of transcription factors and their expression aberrancies could result in tumor development 789.
Altogether, since P21 and P27 have crucial roles in cell cycle regulation, it is likely that leukemic cells profit from changes in their gene expression. Since few studies have been conducted to evaluate the importance of such proteins in ALL patients, we therefore investigated the expression patterns of pivotal cell cycle genes (P21 and P27) in newly diagnosed ALL cases.
Methods
Patients and healthy people
In the present study, we assessed 38 bone marrow (BM) and 14 peripheral blood (PB) samples from newly diagnosed ALL patients, and 13 BM and PB samples from healthy individuals. Patient consent was obtained, according to the institutional ethics guidelines for the study (IR.SBMU.RETHECH.REC.1396.800). Diagnosis of ALL was confirmed by morphology characteristics, immunophenotyping, and molecular analysis. For collection of control samples, we utilized BM and PB specimens from patients with no hematologic malignancies and whom were referred to the lab for routine checkup by physicians.
RNA extraction and cDNA synthesis
Total cellular RNA was extracted using RNeasy Kit (Qiagen, Germany). The integrity of RNA was measured by the NanoDrop (Thermo Scientific, Wilmington, North Carolina, USA). High purity (OD 260/280 nm ratio >1.8) was observed for all samples. Subsequently, cDNA was synthesized from 1 μg of RNA to a final volume of 20 μL by means of a cDNA Synthesis Kit (ThermoFisher Scientific, Waltham, MA, USA). An equal amount of cDNA from controls and patients (50 ng of cDNA) was used as a substrate for qRT- PCR amplification.
Real-Time PCR (qRT-PCR)
The primers used in the present study were designed via Oligo 7.56 software and the data were checked in the NCBI Blast database. The primer sequences were:
P27 forward (5- AGCAATGCGCAGGAATAAGGAAGC-3) and
P27 reverse (5- CACAGAACCGGCATTTGGGGAACC -3),
P21 forward (5- ACTAGGCGGTTGAATGAGAG -3) and
P21 reverse (5- GAGAGGAAAAGGAGAACACG-3), and
ABL forward (5-AGTCTCAGGATGCAGGTGCT-3) and
ABL reverse (5-TAGGCTGGGGCTTTTTGTAA),
according to previous studies; ABL was selected as a housekeeping gene in this study 10.
Consequently, P21, P27 and ABL mRNA expression in the patient and healthy volunteer samples were analyzed by qRT-PCR (Rotor-Gene 6000, Qiagen, Germany). The components of the qRT-PCR reaction for each target consisted of 1 μL of template target cDNA, 1 μL of forward and reverse primer, 7 μL of Real Plus 2x Master Mix Green-Low ROX (Ampliqon, Denmark), and 6 μL water for a total reaction volume of 15 μL. For each qRT-PCR reaction, a standard curve was produced, using five consecutive 1:10 dilutions of a positive sample (1, 0.1, 0.01, and 0.001). The thermal cycler program for each reaction included an initial hold at 95°C for 10 minutes, followed by 40 cycles of denaturation at 95°C for 10 seconds and annealing/extension at 60°C for 30 seconds. The relative quantification of mRNA expression for each sample (fold change = FQ) was calculated using the Livak method (2-ΔΔCT) 11.
Statistical Analysis
The SPSS statistics software (version 16.0) and Graph Pad Prism software (version 6.07) were used for data analysis. In SPSS, LOG was used for normalizing distribution of (2-ΔΔCT) for performing statistical analyses. The Shapiro-Wilk test was applied to determine the normal distribution of the data. One-way ANOVA or Kruskal-Wallis for multi-state variables and t-test or Mann-Whitney U test for two state variables were used according to the Shapiro-Wilk test. Pearson’s chi-square test was performed to analyze correlations. Two-tailed P<0.05 was considered to be significant.
Results
Specification of patient samples
In this study, the expression levels of P21 and P27 were analyzed in peripheral blood and bone marrow specimens collected from 52 patients with de novo ALL. Samples were of different age, sex, and distinct malignant morphological characteristics Table 1. Patient prevalence was determined according to immunophenotype classification, including 3 pro B-ALL, 11 early pre-B-ALL, 9 pre B-ALL, 4 mature B-ALL, and 11 T-ALL.
| Study population | |
| Age (years) | 1-89 |
| Male Sex Female | 30 (57.70%)22 (42.30%) |
| Peripheral blood Sample type Bone marrow | 14 (26.92%)38 (73.08%) |
| Range Blast percent | 20-98 |
P21 and P27 expression in ALL patients and normal controls
Our results revealed that P21 and P27 expression were decreased in ALL patients in comparison to the normal control group (8.33- and 1.69-fold change, respectively) Figure 1.

Differential expression of P21 and P27 according to ALL immunophenotype groups
The differential expression of P21 and P27 between the different immunophenotype groups in ALL patient samples were evaluated by ANOVA test. Statistical analysis of P21 expression showed a significant difference between T-ALL and early pre B-ALL (P=0.03). However, P27 expression was not significantly different among the immunophenotype groups (P= 0.35) Figure 2.


P21 and P27 expression in B-ALL patients and control group
P21 and P27 expression in B-ALL and control group were evaluated using t-test. P21 expression in the B-ALL group was significantly different that of the control group (P=0.001). P27 expression was not significantly different between the B-ALL group and the control group (P=0.47) Figure 3.
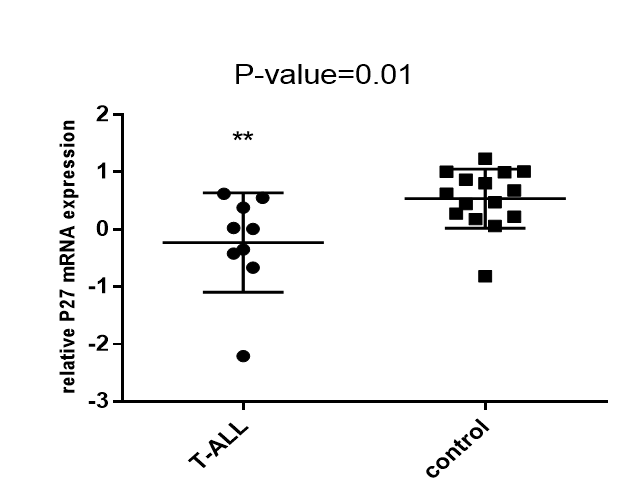

P21 and P27 expression in T-ALL patient group and control group
P21 and P27 expression in T-ALL group and control group were evaluated using t-test. There appeared to be a significant difference between the T-ALL patients and controls Figure 4.
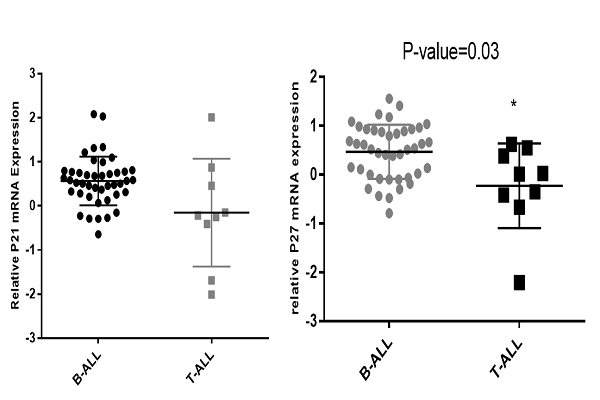
Expression of P21 and P27 in T-ALL and B-ALL patients
The differential expression of P21 and P27 in the T-ALL patient samples versus B-ALL patient samples was evaluated by t-test. Statistical analysis of P21 and P27 expression showed a significant difference between T-ALL and B-ALL patients (P=0.03) Figure 5.
Discussion
Genetic abnormalities including gene mutations and trans-locations are insufficient to generate the whole leukemia characteristics in most patients. Indeed, both genetic and epigenetic modifications induce cooperating altered cell cycle progression and inhibition of apoptosis, which are required for establishment of neoplasms 121314. As an intricate process, cell cycle regulation can undergo different alterations resulting in cell proliferation abnormalities and cancer progression. Proper cell division is ensured by various mechanisms controlling cell cycle 1516. These mechanisms include CDK regulation through cyclins, CDK inhibitors, and phosphorylation 17. Studies have revealed that the expression of cell cycle regulators, such as P21 and P27, is relevant to aggressive tumor patterns in human malignancies 1819202122. Although cell cycle deregulation have been indicated in different types of cancers, there are insufficient studies evaluating the expression patterns of cell cycle regulators in ALL with variable outcomes. Therefore, we evaluated the expression of P21 and P27 as two main CDKIs involved in cell cycle regulation.
In the present study, we found that P21 and P27 expression levels were diminished by 8.33- and 1.69-fold, respectively, in ALL patient samples compared to the control group. Our results demonstrated a decreased expression of P21 in 94% and reduced expression of P27 in 50% of the ALL patients in our study.
In agreement with our results, Roman-Gomez et al. (2002) has evaluated the P21 promoter methylation pattern in ALL patients; P21 hypermethylation was observed in 41% of their patients and was strongly correlated with decreased expression of P21 in cancerous cells 5. Pellikainen et al. (2003) demonstrated a decreased expression of P21 in their study on breast cancer samples (by IHC techniques), which was concordant with our results 23. Radosevic et al. (2001) also detected very low levels of P21 protein in their fresh AML samples 24. In agreement with our results, Zolota et al. detected P21 only in a small percentage of their BM samples. However, P27 expression was observed in a high percentage of their cases, which is in contrast with our findings 25. It seems that decreased expression of P21 as a tumor suppressor gene in leukemic cells is the result of activation of oncogenic pathways, and facilitates leukemic growth.
In a study by Letestu et al. (2004), P27 expression was evaluated in Mantle cell lymphoma (MCL) using western blot technique. The authors observed an absence of P27 expression in 72% of their patients while P27 median expression was observed in only 28% of patients. They also showed that survival of P27 negative patients was greater than that of P27 positive patients 26. Møller et al. (1999) investigated P27Kip1 expression in non-Hodgkin’s lymphoma (NHL) patients by immunohistochemistry. They demonstrated that lower expression of P27Kip1 was predictive of poor survival in indolent and aggressive NHL 27. Chiarle et al. (2000) also investigated the expression of P21 and P27 in MCL. They observed loss of P27 in a high percentage of MCL cases due to proteasome-mediated degradation; the loss of P27 was associated with poor outcomes 28. Halvorsen et al. (2003) also evaluated the expression of P27 and phosphatase and tensin homolog (PTEN) in primary prostatic carcinomas. In their study, P27 low expression was associated with high clinical stage of prostatic carcinoma 29.
Moreover, in an epigenetic analysis by Chim et al. (2005) and Takeuchi et al. (2011) on cell cycle regulators, such as P21 and P27, in patients with acute leukemia, P21 hypermethylation was not detected and P27 methylation was rarely demonstrated 430. Winkler et al. (2010) also reported P27 overexpression in patients with CLL, which was associated with higher risk groups 31. Van de Putte et al. (2002) showed that P21 expression was increased in 20% of patients with early squamous cervical cancer, while P27 expression was decreased in 80% of their samples 32. In malignant ovarian tumors, Lee et al. (2011) showed P21 overexpression compared with borderline tumors. P27 expression was not significantly changed among benign, borderline, and malignant tumors in histologic evaluations of ovarian tumors. The authors showed that cytoplasmic P27Kip1 was relevant and associated with shorter overall survival in ovarian carcinomas 33.
The discrepancies among the reported studies possibly arise from the employment of different analysis methods, from protein degradation, and/or from unexpected functions. P21 and P27 are believed to exert their inhibitory functions through similar pathways as a result of high homology in their primary structures 34. It seems that P21 and P27 genes are subject to changes at the transcriptional level in ALL patients. Generally, P21 and P27 can be considered as strong tumor suppressors in ALL leukemic cells; their expression can, indeed, be influenced by epigenetic modifications or alternative mechanisms. It is postulated that P21 and P27 tightly regulate cell cycle such that their absence or aberration can result in proliferation and tumor growth in ALL. Moreover, the appropriate cellular localization of P21 and P27 can govern their tumor suppressor functions 353637.
In conclusion, limited studies are available concerning expression, regulation, and localization patterns of cell cycle regulators, such as P21 and P27, in hematologic malignancies. Future studies should be done to assess these genes not only at the transcription level but also at the post-transcriptional and translational levels. Evaluation of expression of these genes can enhance our knowledge about the molecular mechanisms involved in ALL formation and progression. Furthermore, these evaluations can help in developing ALL risk classifications precisely and in yielding more effective therapies for ALL patients.
Conclusions
Since P21 and P27 are able to influence the activity of both cyclins and CDKs, the decreased expression of these genes reduces P21- and P27- mediated suppressive effects on cyclins and CDKs. These events can, therefore, facilitate the activation of cyclins and CDKs, which could result in cancer progression in ALL patients.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License (CCBY4.0) which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
List of abbreviations
ALL: Acute Lymphoblastic Leukemia; CDK: Cyclin-Dependent Kinases; CDKI: Cyclin-Dependent Kinase Inhibitor; CIP/KIP: CDK interacting protein/Kinase inhibitory protein ; CLL: chronic lymphocytic leukemia; INK4: Inhibitors of CDK4; MCL: Mantle cell lymphoma; NHL: non-Hodgkin’s lymphoma; QPCR: quantitative polymerase chain reaction
Ethics approval and consent to participate
Patient consent was obtained, according to the institutional ethics guidelines for the study (IR.SBMU.RETHECH.REC.1396.800)
Competing interests
The authors declare no conflict of interest.
Funding
Shahid Beheshti University of Medical Sciences, Shahid Beheshti, Iran.
Authors' contributions
All authors contributed to the design of the research. MKH, MHM, PKH and SL collected the
Data. HA, AGH and MA conducted analysis and interpretation of data. All authors drafted
The first version. MKH, MHM, PKH, SL, HA and ZKH edited the first draft. All authors reviewed, Commented and approved the final draft.
Acknowledgments
We would like to express our gratitude to the department of hematology and blood banking, Shahid Beheshti University of Medical Sciences and HSCT Research Center of Taleghani hospital.
References
-
Yahya
Raida S,
Fouda
Manal I,
El-Baz
Hatim A,
Mosa
Tamer E,
ElMaksoud
Mohamed D Abd.
Serum survivin and tp53 gene expression in children with acute lymphoblastic leukemia. Iranian journal of public health.
2012;
41
:
37
.
-
Jaime-Pérez
José Carlos,
Pinzón-Uresti
Mónica Andrea,
Jiménez-Castillo
Raúl Alberto,
Colunga-Pedraza
Julia Esther,
González-Llano
Óscar,
Gómez-Almaguer
David.
Relapse of childhood acute lymphoblastic leukemia and outcomes at a reference center in Latin America: organomegaly at diagnosis is a significant clinical predictor. Hematology.
2018;
23
:
1-9
.
-
Chokkalingam
Anand P,
Bartley
Karen,
Wiemels
Joseph L,
Metayer
Catherine,
Barcellos
Lisa F,
Hansen
Helen M,
Aldrich
Melinda C,
Guha
Neela,
Urayama
Kevin Y,
Scélo
Ghislaine.
Haplotypes of DNA repair and cell cycle control genes, X-ray exposure, and risk of childhood acute lymphoblastic leukemia. Cancer Causes & Control.
2011;
22
:
1721
.
-
Chim
CS,
Wong
ASY,
Kwong
YL.
Epigenetic inactivation of the CIP/KIP cell‐cycle control pathway in acute leukemias. American journal of hematology.
2005;
80
:
282-287
.
-
Roman-Gomez
Jose,
Castillejo
Juan Antonio,
Jimenez
Antonio,
Gonzalez
Maria Gracia,
Moreno
Fernanda,
del Carmen Rodriguez
Maria,
Barrios
Manuel,
Maldonado
Juan,
Torres
Antonio.
5′ CpG island hypermethylation is associated with transcriptional silencing of the p21CIP1/WAF1/SDI1 gene and confers poor prognosis in acute lymphoblastic leukemia. Blood.
2002;
99
:
2291-2296
.
-
Barata
Joao T,
Cardoso
Angelo A,
Nadler
Lee M,
Boussiotis
Vassiliki A.
Interleukin-7 promotes survival and cell cycle progression of T-cell acute lymphoblastic leukemia cells by down-regulating the cyclin-dependent kinase inhibitor p27kip1. Blood.
2001;
98
:
1524-1531
.
-
Kitaura
Hirotake,
Shinshi
Minako,
Uchikoshi
Yasuko,
Ono
Takashi,
Tsurimoto
Toshiki,
Yoshikawa
Hiroshi,
Iguchi-Ariga
Sanae MM,
Ariga
Hiroyoshi.
Reciprocal regulation via protein-protein interaction between c-Myc and p21 cip1/waf1/sdi1 in DNA replication and transcription. Journal of Biological Chemistry.
2000;
275
:
10477-10483
.
-
Coqueret
Olivier,
Gascan
Hugues.
Functional interaction of STAT3 transcription factor with the cell cycle inhibitor p21 WAF1/CIP1/SDI1. Journal of Biological Chemistry.
2000;
275
:
18794-18800
.
-
Harris
Thurl E,
Albrecht
Jeffrey H,
Nakanishi
Makoto,
Darlington
Gretchen J.
CCAAT/enhancer-binding protein-α cooperates with p21 to inhibit cyclin-dependent kinase-2 activity and induces growth arrest independent of DNA binding. Journal of Biological Chemistry.
2001;
276
:
29200-29209
.
-
Abraham
Ajay,
Varatharajan
Savitha,
Karathedath
Sreeja,
kumar Jeyavelu
Ashok,
Velayudhan
Shaji R,
Ahmed
Rayaz,
Abraham
Aby,
George
Biju,
Chandy
Mammen,
Srivastava
Alok.
ABC Transporter Expression in Acute Myeloid Leukemia: Association with in Vitro Cytotoxicity and Prognostic Markers. 2012
.
-
Livak
Kenneth J,
Schmittgen
Thomas D.
Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods.
2001;
25
:
402-408
.
-
Mullighan
Charles G,
Goorha
Salil,
Radtke
Ina,
Miller
Christopher B,
Coustan-Smith
Elaine,
Dalton
James D,
Girtman
Kevin,
Mathew
Susan,
Ma
Jing,
Pounds
Stanley B.
Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature.
2007;
446
:
758
.
-
Shahreza
Fatemeh Dehghan.
From oxidative stress to endothelial cell dysfunction.. J Prev Epidemiol..
2016;
1
(1)
:
e04
.
-
Mousavi Movahhed SM
Beladi Mousavi SS, Hayati F, Shayanpour S, Halili SA, Sabetnia L, Khazaei Z..
The relationship between chronic kidney disease and cancer.. J Nephropathol..
2018;
7
:
115-16
.
-
Salarpour
Fatemeh,
Goudarzipour
Kourosh,
Mohammadi
Mohammad Hossein,
Ahmadzadeh
Ahmad,
Faraahi
Sara,
Farsani
Mehdi Allahbakhshian.
Evaluation of CCAAT/Enhancer Binding Protein (C/EBP) Alpha (CEBPA) and Runt‐Related Transcription Factor 1 (RUNX1) Expression in Patients with De Novo Acute Myeloid Leukemia. Annals of human genetics.
2017;
81
:
276-283
.
-
Goudarzipour
Kourosh,
Ahmadzadeh
Ahmad,
Mohammadi
Mohammad Hossein.
Changes of AML 1 and P53 tumor suppressor gene expression in patients de novo acute myeloid leukemia. Journal of Paramedical Sciences.
2017;
8
:
39-45
.
-
Baekelandt
Mark,
Holm
Ruth,
Tropé
Claes G,
Nesland
Jahn M,
Kristensen
Gunnar B.
Lack of independent prognostic significance of p21 and p27 expression in advanced ovarian cancer: an immunohistochemical study. Clinical cancer research.
1999;
5
:
2848-2853
.
-
Taguchi
Ken‐Ichi,
Aishima
Shin‐Ichi,
Asayama
Yoshiki,
Kajiyama
Kiyoshi,
Kinukawa
Naoko,
Shimada
Mitsuo,
Sugimachi
Keizo,
Tsuneyoshi
Masazumi.
The role of p27kip1 protein expression on the biological behavior of intrahepatic cholangiocarcinoma. Hepatology.
2001;
33
:
1118-1123
.
-
Fiorentino
Michelangelo,
Altimari
Annalisa,
D’Errico
Antonia,
Gabusi
Elena,
Chieco
Pasquale,
Masetti
Michele,
Grigioni
Walter Franco.
Low p27 expression is an independent predictor of survival for patients with either hilar or peripheral intrahepatic cholangiocarcinoma. Clinical cancer research.
2001;
7
:
3994-3999
.
-
Kang
Yun Kyung,
Kim
Woo Ho,
Jang
Ja June.
Expression of G1-S modulators (p53, p16, p27, cyclin D1, Rb) and Smad4/Dpc4 in intrahepatic cholangiocarcinoma. Human pathology.
2002;
33
:
877-883
.
-
Bukholm
Ida K,
Nesland
Jahn M.
Protein expression of p53, p21 (WAF1/CIP1), bcl-2, Bax, cyclin D1 and pRb in human colon carcinomas. Virchows Archiv.
2000;
436
:
224-228
.
-
M
Bahadoram S, Bahadoram M, Barahman M, Khazaei Z, Amiri M. Davoodi.
Impact of cancers on the kidney function and structure; an ignored entity.. J Renal Inj Prev..
2018;
7
:
112-115
.
-
Pellikainen
MJ,
Pekola
TT,
Ropponen
KM,
Kataja
VV,
Kellokoski
JK,
Eskelinen
MJ,
Kosma
VM.
p21WAF1 expression in invasive breast cancer and its association with p53, AP-2, cell proliferation, and prognosis. Journal of clinical pathology.
2003;
56
:
214-220
.
-
Radosevic
N,
Delmer
A,
Tang
R,
Marie
JP,
Ajchenbaum-Cymbalista
F.
Cell cycle regulatory protein expression in fresh acute myeloid leukemia cells and after drug exposure. Leukemia.
2001;
15
:
559
.
-
Zolota
Vassiliki,
Sirinian
Chaido,
Melachrinou
Maria,
Symeonidis
Argyrios,
Bonikos
Dionysios S.
Expression of the regulatory cell cycle proteins p21, p27, p14, p16, p53, mdm2, and cyclin E in bone marrow biopsies with acute myeloid leukemia. Correlation with patients’ survival. Pathology-Research and Practice.
2007;
203
:
199-207
.
-
Letestu
R,
Ugo
V,
Valensi
F,
Radford-Weiss
I,
Nataf
J,
Levy
V,
Gribben
JG,
Troussard
X,
Ajchenbaum-Cymbalista
F.
Prognostic impact of p27KIP1 expression in cyclin D1 positive lymphoproliferative disorders. Leukemia.
2004;
18
:
953
.
-
Møller
Michael B,
Skjødt
Karsten,
Mortensen
Leif S,
Pedersen
Niels T.
Clinical significance of cyclin‐dependent kinase inhibitor p27Kip1 expression and proliferation in non‐Hodgkin\'s lymphoma: independent prognostic value of p27Kip1. British journal of haematology.
1999;
105
:
730-736
.
-
Chiarle
Roberto,
Budel
Leo M,
Skolnik
Jeffrey,
Frizzera
Glauco,
Chilosi
Marco,
Corato
Alessandra,
Pizzolo
Gianni,
Magidson
Jory,
Montagnoli
Alessia,
Pagano
Michele.
Increased proteasome degradation of cyclin-dependent kinase inhibitor p27 is associated with a decreased overall survival in mantle cell lymphoma. Blood.
2000;
95
:
619-626
.
-
Halvorsen
Ole J,
Haukaas
Svein A,
Akslen
Lars A.
Combined loss of PTEN and p27 expression is associated with tumor cell proliferation by Ki-67 and increased risk of recurrent disease in localized prostate cancer. Clinical cancer research.
2003;
9
:
1474-1479
.
-
Takeuchi
Seisho,
Matsushita
Masahide,
Zimmermann
Martin,
Ikezoe
Takayuki,
Komatsu
Naoki,
Seriu
Taku,
Schrappe
Martin,
Bartram
Claus R,
Koeffler
H Phillip.
Clinical significance of aberrant DNA methylation in childhood acute lymphoblastic leukemia. Leukemia research.
2011;
35
:
1345-1349
.
-
Winkler
Dirk,
Schneider
Christof,
Zucknick
Manuela,
Bogelein
Daniela,
Schulze
Kerstin,
Zenz
Thorsten,
Mohr
Julia,
Philippen
Angela,
Huber
Henriette,
Buhler
Andreas.
Protein expression analysis of chronic lymphocytic leukemia defines the effect of genetic aberrations and uncovers a correlation of CDK4, P27 and P53 with hierarchical risk. haematologica.
2010;
:
haematol. 2010.025734
.
-
Van de Putte
Gregg,
Holm
Ruth,
Lie
A Kathrine,
Tropé
Claes G,
Kristensen
Gunnar B.
Expression of p27, p21, and p16 protein in early squamous cervical cancer and its relation to prognosis. Gynecologic oncology.
2003;
89
:
140-147
.
-
Lee
Yoon Hee,
Heo
Jin-hyung,
Kim
Tae Heon,
Kang
Haeyoun,
Kim
Gwangil,
Kim
Jiyoung,
Cho
Sang-ho,
An
Hee Jung.
Significance of cell cycle regulatory proteins as malignant and prognostic biomarkers in ovarian epithelial tumors. International Journal of Gynecological Pathology.
2011;
30
:
205-217
.
-
Russo
Alicia A,
Jeffrey
Philip D,
Patten
Andrea K,
Massagué
Joan,
Pavletich
Nikola P.
Crystal structure of the p27Kip1 cyclin-dependent-kinase inibitor bound to the cyclin A–Cdk2 complex. Nature.
1996;
382
:
325
.
-
Cheng
Mangeng,
Olivier
Paul,
Diehl
J Alan,
Fero
Matthew,
Roussel
Martine F,
Roberts
James M,
Sherr
Charles J.
The p21Cip1 and p27Kip1 CDK ‘inhibitors’ are essential activators of cyclin D‐dependent kinases in murine fibroblasts. The EMBO journal.
1999;
18
:
1571-1583
.
-
Alt
Jodi R,
Gladden
Andrew B,
Diehl
J Alan.
p21Cip1 promotes cyclin D1 nuclear accumulation via direct inhibition of nuclear export. Journal of Biological Chemistry.
2002;
277
:
8517-8523
.
-
Müller
Daniel,
Thieke
Katja,
Bürgin
Andrea,
Dickmanns
Achim,
Eilers
Martin.
Cyclin E‐mediated elimination of p27 requires its interaction with the nuclear pore‐associated protein mNPAP60. The EMBO journal.
2000;
19
:
2168-2180
.
Comments
Downloads
Article Details
Volume & Issue : Vol 5 No 7 (2018)
Page No.: 2518-2527
Published on: 2018-07-28
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
- HTML viewed - 10440 times
- Download PDF downloaded - 2085 times
- View Article downloaded - 0 times
