

Copyrights: Nadia H. Ismaeil, Amany A. Osman, Elham H.A. Ali, Laila A. Rashed, Manal A. Saleh, 2017. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Introduction: The present work aims to examine the possible role of stem cells on biochemical markers and histopathological alterations of hypoxia caused by sodium nitrite (NaNO2) toxicity in testes of male rats.
Methods: In this study, 96 adult male albino rats were divided into 6 groups (16 rats each). Group 1 (G1) was the control group and received distilled H2O. Group 2 (G2) received daily NaNO2 (35 m/kg bwt/ day) via subcutaneous injection for 3 weeks. Group 3 (G3) received NaNO2 for 2 weeks and were then injected once with 2*106 mesenchymal stem cells (MSCs) intravenously and sacrificed 4 weeks later. Group 4 (G4) received NaNO2 for 2 weeks and were then injected with 2*106 MSCs followed by daily NaNO2 injection for 1 week; rats in G4 were sacrificed 4 weeks from MSCs treatment. Group 5 (G5) rats were treated with NaNO2 for 2 weeks and then left to recover for 4 weeks. Finally, Group 6 (G6) rats were treated with NaNO2 for 3 weeks and left to recover for 3 weeks, after which point they were sacrificed.
Results: The results showed that NaNO2 caused oxidative damage and histopathological alterations in the rat testes, as well as increased the levels of testes malondialdehyde (MDA), nitric oxide (NO) and DNA fragmentation percentage (DNA F %). Moreover, NaNO2 decreased the elevated activities of testes catalase (CAT) and total antioxidant activity (TAA), in comparison to the control group. The histological results illustrated different distortions, vacuolization and lipid accumulations in interlobular space as well as diminution of inter cellular germ cell layers, absence of Leydig cells, irregular basement membrane of tubule, and separation within spermatogenic cells. In addition, congestion and dilation of intertubular and peripheral blood capillaries were found. Nevertheless, the administration of stem cells reduced the danger actions of sodium nitrite by enhancing biochemical marker concentration.
Conclusion: There was an improvement in the histology of the rat testes, including a relatively normal order in the different stages of spermatogonia and loss of different stages of spermatocytes. Regarding the recovery period, there was also a significant improvement in each of the biochemical parameters assessed and in the histological lesions.
Introduction
Sodium nitrite (NaNO2) is a familiar preservative for meat and dye with both dangerous and beneficial effects. Processed meats, such as hot dogs, bacon, sliced deli meats and sausage, contain NaNO2 in its list constituents. Sodium nitrite is utilized occasionally for medicinal purposes and may be employed as a component in the treatment of sickle-cell anemia and heart attack Cosby et al., 2003Mack et al., 2008. It is a water soluble and inorganic salt extensively used in different industries, including the agricultural, textile processing, chemical, coloring, and disinfectant industries U.S.DHHS, 2001.
Hypoxia refers to low oxygen concentration in the body or organs. The condition begins from an inequality between the quantity of oxygen needed through the body and the quantity of oxygenated blood that is provided Maher et al., 2008.
Numerous studies have shown that the harmful impact of NaNO2 induced hypoxia is related to inflammation, oxidative stress and methemoglobinemia, which induce injury and dysfunction of different organs Al-Gayyar et al., 2014El-Sheikh and Khalil, 2011Hassan et al., 2009Salama et al., 2013.
The body responds to hypoxia by adaptive reactions, such as angiogenesis, smooth muscle relaxation and vasodilatation, thereby elevating tissue blood supply and compensating for the lack of oxygen. The hypoxia-driven adaptive reactions also cause an elevation in the temperature of the testes Farias et al., 2005. The rise in temperature activates apoptosis in cells of the seminiferous tubules of the testis. The exposure to continuous and intermittent hypoxia changes the male mouse reproductive system. Greater testicular damage have been observed under continuous hypoxia. With intermittent hypoxia, lesions are less essential, suggesting that cycles of normoxia compensate the effects on the testicle Vargas et al., 2011.
Stem cells are able to regenerate and are capable of differentiating into specialized cell types Bruder et al., 1997Chopp et al., 2000El Asmar et al., 2011Sato et al., 2005. Anderson et al. (2001) reported that stem cells have the potential to treat various diseases, inducing diabetes, liver, heart illness and infertility. Adult mesenchymal stem cells (MSCs) are believed to be highly promising applicant cells for renewal applications for the reason that they have a high proliferative capability and the ability to differentiate into specified cells Caterson et al., 2002Nöth et al., 2002. In fact, MSCs are self-renewing cells which are able to differentiate into several mesodermal tissues, e.g. bone, fat, cartilage and muscle Pittenger et al., 1999, and to migrate toward damaged tissue sites Fujimoto et al., 2012. The present work aims to evaluate the action of NaNO2 induced hypoxia on male rat testes and the therapeutic role MSCs play in the deleterious effects of NaNO2 on testes.
Materials - Methods
Experimental animals
In the present study 96 adult male albino rats (weighing 150-180 g) were used. Rats were obtained from Ain Shams Hospital Animal House. Rats were left for two weeks for acclimatization before starting the experiments. Animal procedures and experimental protocols were approved by Ain Shams University authorities; they were in accordance with the Egyptian animal protection rules and consistent with the guidelines of the European Communities (EC) (1986).
Sodium nitrite (NaNO2) administration
NaNO2 (7632-00-0) was obtained from Alahram Company (Al-Sadat city, Egypt) and dissolved in distilled water at 27°C. It was administered subcutaneously to rats at a dose of 35 mg/kg bwt/day, as previously described by Bhanumathy et al. (2010) Bhanumathy et al., 2010.
Isolation, propagation, identification and labeling of MSCs derived from bone marrow of rats
Bone marrow was removed from the tibiae and femurs of male rats, aged 6 weeks old, and flushed in Dulbecco’s modified Eagle’s medium (DMEM, GIBCO/BRL) enhanced with 10% fetal bovine serum (FBS, GIBCO/BRL). The nucleated cells were then separated in a density gradient via Ficoll/Paque (Pharmacia) and re-suspended in whole culture medium complemented with 1% penicillin streptomycin (GIBCO/BRL). Cells were cultured at 37°C, 5% humidified CO2 for 12-14 days during the primary culture. The medium were replaced each 2-3 days. When colonies grew large (80-90% confluence), the culture was rinsed with PBS twice and the cells were trypsinized with 0.25% trypsin in 1 mM EDTA (GIBCO/BRL) for 5 min at 37°C. After centrifugation, cells were re-suspended with serum-supplemented culture and kept warm in 50cm2 culture flasks (Falcon). The resultant media were considered first route supernatant cultures Alhadlaq and Mao, 2004. On day 14, the adherent cell colonies were trypsinized and counted.
Cells were recognized as MSCs by the identification of their morphology, adherence, and ability to transform into osteocytes Jaiswal et al., 1997 and chondrocytes Seo et al., 2009. Transformation into osteocytes was attained by adding 1 to 1000 nM dexamethasone, 0.25 mM ascorbic acid, and 1 to 10 mM beta-glycerophosphate to the culture. Transformation of MSCs to osteoblasts was attained through morphological changes and Alzarin red staining of differentiated osteoblasts. Transformation to chondrocytes was attained by adding 500 ng/ml bone morphogenetic protein-2 (BMP-2; R&D Systems) and 10 ng/ml transforming growth factor-β3 (TGF- β3) (Peprotech, London) for 3 weeks Seo et al., 2009. In vitro transformation into chondrocytes was verified by morphological changes and Alcian blue staining of differentiated chondrocytes. Flow cytometric analysis of cultured MSCs surface markers was also done.
In the current study, MSCs were tagged with PKH26 (Sigma, Saint Louis, MO), consistent with the manufacturer’s advice. Cells were intravenously injected once into the rat tail vein at a dose of 2*106 cells, according to previously published by Kebriaei et al. (2009) Kebriaei et al., 2009.
Experimental Design
Rats were randomly split into six main groups with 16 rats each. Group 1 (G1) received distilled water and served as the control. Group 2 (G2) received daily subcutaneous (s.c.) NaNO2 injections at a dose of 35 mg/kg bwt/ day for 3 weeks. Group 3 (G3) received NaNO2 for 2 weeks and were then injected once with MSCs (2*106 cells) intravenously; rat were sacrificed 4 weeks after MSC injection. Group 4 (G4) were treated with NaNO2 for 2 weeks and then treated with MSCs at 1 week following NaNO2 injection; rats were sacrificed 4 weeks from MSC injection. Group 5 (G5) were treated with NaNO2 for 2 weeks and then left to recover for 4 weeks and then sacrificed. Lastly, Group 6 (G6) received NaNO2 for 3 weeks then left to recover for 3 weeks and then sacrificed.
Biochemical Studies
Testes samples were prepared by taking a weighted part of the testes and homogenizing in the recommended saline. Biochemical analyses were conducted, which included colorimetric determination of NO (carried out according to Miranda et al. (2001) Miranda et al., 2001 using modified Griss reagent), measurement of malondialdehyde in the tissue (using a modified thiobarbituric acid (TBA) assay consistent with Draper and Hadley (1990) Draper and Hadley, 1990), percentage of DNA fragmentation (as determined by quantitative analysis using diphenylamine assay, according to the method described by Sharawy (2013) Sharawy, 2013, and catalase activity determination (through a method used by Bock and Pavelka (1980) Bock et al., 1980).
Moreover, total antioxidant activity was verified, according to Koracevic et al. (2001) Koracevic et al., 2001.
Histopathological Examination
In the present study, testes specimens were carefully dissected and fixed in Bouin’s solution for routine histological analysis, and implanted in paraffin wax at 60oC. Serial transverse sections were then cut at 5-6 microns in thickness using Cambridge Rocking Microtome and affixed to slides. For general histological examination, sections were stained with Hematoxylin and Eosin (Drury and Wallington, 1980).
Statistical Analysis
Reported data represented the mean ± SE for 8 animals per group. For statistical analysis, one-way analysis of variance (ANOVA) and post-HOC test ("least significant difference (LSD) analysis) were completed using statistical package for social science (SPSS) for Windows software (version 17). Statistical significance was set at p<0.05.
Results
Biochemical investigation
The results of the biochemical parameters are shown in Table 1 . Oxidative stress parameters showed a significant increase in all treated groups with regards to the following: the value of testes nitric oxide (NO) contents, malondialdehyde (MDA) contents, and DNA fragmentation percentage (DNA F%). On the other hand, there was a significant reduction in catalase activity (CAT) and total antioxidant activity (TAA) of G2 group as compared to the G1 control group. Only G3 rats showed a non-significant change in MDA contents.
Moreover, there was a significant reduction in the recovery groups and the MSC treated groups with respect to testes NO contents, MDA contents, and DNA F%, when compared with those rats in G2 group. Conversely, testes catalase activities (CAT) and total antioxidant activities (TAA) were significantly increased in stem cell groups and recovery groups, compared to G2 rats.
Histological investigations
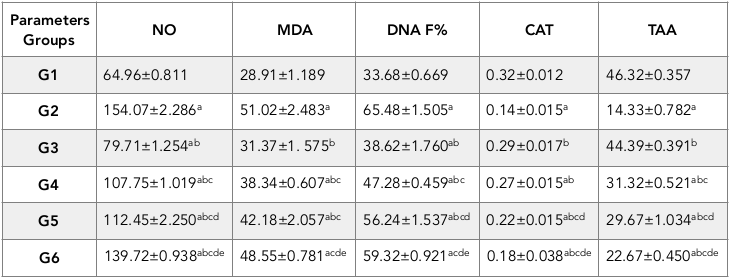
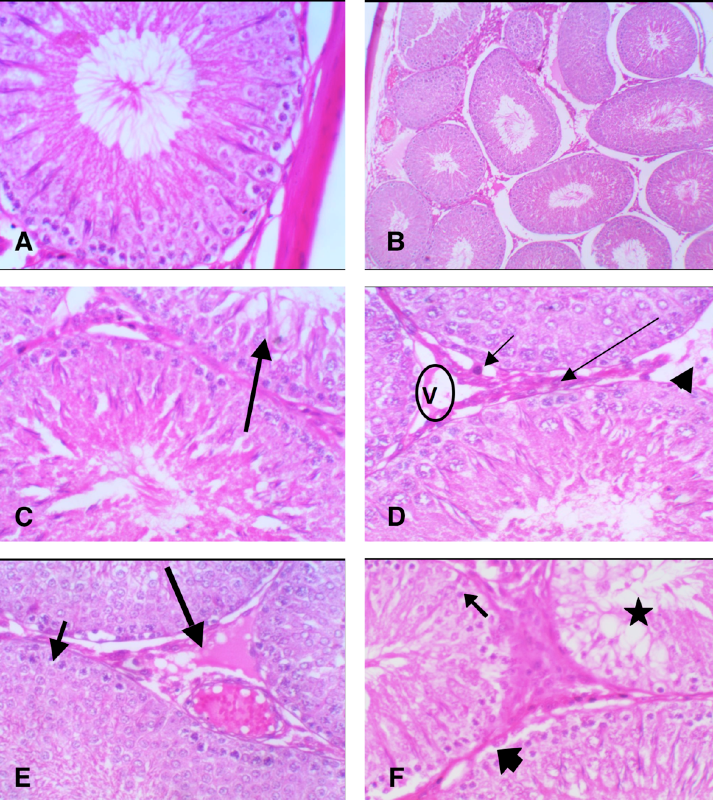
In the present study, sections from the testes of the control rats (G1) showed normal structures of seminiferous tubules and interstitial tissue ( Figure 1A ).
Spermatogonia was normal too, with primary and secondary spermatocytes/spermatids developing as standard, and moving towards Sertoli cells ( Figure 1B ).
The hypoxic group (G2) showed seriously injured seminiferous tubules. Microscopic alterations were regarded as characteristic indications for seriously injured tubules. The greater part of the seminiferous tubules showed vacuolization, lipid accumulation in intertubular space with gelatinous material in intertubular connective tissues, and diminution of Leydig cells. Also, germ cell layers were reduced ( Figure 1C ). Abnormally shaped seminiferous tubules manifested irregular tubule basement membranes within necrotic areas of the spermatogenic cells layers ( Figure 1D ). Meanwhile, some seminiferous tubules appeared with cellular debris in the tubular lumen ( Figure 1E ). Numerous giant cells were observed with cluster multinucleated nesting arrangement, karyorrhexis of Leydig cells, depletion of spermatocytes, hyaline material with absence of Leydig cells in interstitial connective tissues, and separated seminiferous tubule basement membranes.
Specifically, the G3 group revealed significant improvement of basement membrane and seminiferous tubules borders; some elongated tubules as shown in Figure 1F . Moreover, the seminiferous tubules showed a nearly normal pattern of all the spermatogenic stages, activated Sertoli cells, and a slightly disordered and thickened outer basal layer.
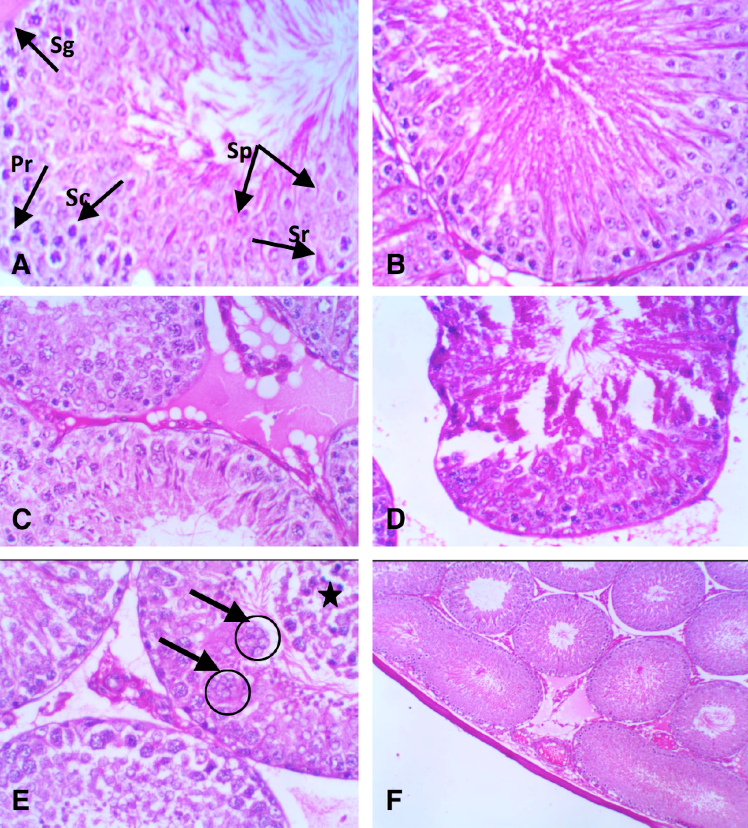
There were definite improvements of histopathological lesions seen after MSC therapy. These are clearly demonstrated in Figure 2A . The testicular structure sections from G4 rats showed a relatively normal pattern of different seminiferous tubules, accompanied by an intertubular space (with dilated and congested interstitial blood vessels), hyaline substances, and a rare disturbance pattern of spermatocytes ( Figure 2B,C ).
The histological structure of sections of the testes of G5 rats ( Figure 2D,E ) illustrated a slight improvement of the testicular structures. They appeared nearly normal, and the seminiferous tubules showed only a few abnormally elongated shapes. The interstitial blood vessels were slightly dilated and congested. There were also Leydig cells with hyaline substances and some degenerative patches in the intertubular spaces. The testes of G6 rats showed a partial amelioration of seminiferous tubules with hyaline material, an absence of Leydig cells in the interstitial spaces, and evident erosion and hydropic degeneration ( Figure 2F ). Recovery of treated G5 rats revealed little pathological change, compared to G6 treated rats.
Discussion
Sodium nitrite is known as an inorganic salt which have a variety of industrial purposes. The adverse health effects of NaNO2 in animals and humans typically lead to the formation of methemoglobin in the blood. This is due to cyanosis which can, at extremely elevated levels, lead to death. People are regularly subjected to sodium nitrite during nutrition and water Pavlova et al., 2013. Hypoxia is recognized to obstruct fertility in man Okumura et al., 2003 and in other mammals. Hypoxia can influence the function of the testes by diminishing the level of testosterone and by disturbing spermatogenesis Farías et al., 2008.
In present study, NaNO2 (at a dose of 35m/kg s.c.) had pathological effects of on the testes of rats. It led to a rise in MDA levels, as compared to control rats (which did not receive NaNO2 daily injections). In accordance with our observation, Vossen and De Smet (2015) Vossen and De Smet, 2015 have shown that antioxidants can be used as therapy to decrease nitrite toxicity Al-Gayyar et al., 2014Al-Gayyar et al., 2016. Interestingly, NO has been connected to an elevation in lipid peroxidation and MDA in both humans
Romeo et al., 2003 and experimental rats Özdamar et al., 2004. A rise in NO following eNOS overexpression has been connected to germ cell apoptosis in a cryptorchidism germ mouse model (i.e. the absence of one or both testes from the scrotum) Ishikawa et al., 2005.
The results obtained from the present work show that hypoxia caused an increase in testes DNA fragmentation percentage after administration of NaNO2. This is compatible with Knaapen et al. (2005) Knaapen et al., 2005 who recorded a rise in DNA fragmentation in adult rats and concluded that NaNO2 induced DNA fragmentation due to oxidative stress. The rigid wrapping of chromatin, which depends on condensation and replacement of histones with protamines, is particularly essential as adult spermatocytes are not capable of fixing DNA damage De Ambrogi et al., 2006Laberge and Boissonneault, 2005. Free radicals possess the capability to precisely injure sperm DNA by assaulting the purine and pyrimidine units and the backbone deoxyribose. Generally, the DNA of sperm is firmly enveloped by protamines shielding it from the assault of free radicals. But, infertile men frequently show a lack of protamination, placing the DNA of the sperm susceptible to ROS assault Oliva, 2006.
On the other hand, free radicals are able to initiate apoptosis in the sperm, causing caspase-mediated enzymatic dilapidation of the sperm DNA Duru et al., 2000Moustafa et al., 2004Villegas et al., 2005Wang et al., 2003. ROS may cause varying degrees of sperm dysfunction, depending on the amount of oxidative stress. Damage from ROS occurs primarily through two routes. First, ROS may be responsible for DNA fragmentation which is usually seen in the spermatozoa of infertile men by causing single- and double-stranded DNA fractures Kodama et al., 1996. Second, higher levels of ROS also may cause damage through a chain of chemical reactions that result in lipid peroxidation of the sperm plasma membrane Alvarez et al., 1987.
The present study clearly demonstrates that there is a considerable rise in DNA fragmentation in testes of NaNO2-treated rats in comparison with control rats. David and Grongnet (2000) showed similar results; they reported that treatment with NaNO2 led to increasing fragmentation David and Grongnet, 2000. More extensive fragmentation of DNA was observed when the time of hypoxia exposure was increased.
In our study, treatment with MSCs may cause MSC differentiation into germ cells in the testis Nayernia et al., 2006. The generation of oxidative stress in infertile rats is reflected by increased production of TBARS, a measure of lipid peroxidation in the homogenates of testes, according to Hussein et al. (2015) Hussein et al., 2015, and which may explain our findings. In the cell membranes, polyunsaturated fatty acid residues react with oxygen radicals normally causing collection of lipid peroxidation yields, several of which also injure proteins and DNA. Leydig cells contain higher membrane lipid matter and may affect the vulnerability of the testes for lipid peroxidation in cadmium treated mice Dobretsov et al., 1977Georgiou et al., 1987Hall, 1994.
Results of our study indicated a reduction in the testes CAT and TAA activities in hypoxic rats compared to control ones. Treatment with MSCs significantly restored those levels. CAT and TAA activities significantly increased in MSC treated rats whereas the NaNO2 group showed a significant decrease in activities compared to the controls. These results could be explained by the inhibition of CAT activity. Titov and Petrenko (2002) suggested that the supraphysiological doses of NaNO2 inhibited catalase activity Titov and Petrenko, 2003. This mechanism could clarify the initial observations that NaNO2 defends H2O2 against damage by catalase activity in hemolysates.
Moreover, applications of 50 – 100 μM NaNO2 hinder myeloperoxidase enzymatic activity, stopping its utilization of H2O2 Knaapen et al., 2005. Treatment with MSCs in the hypoxic groups led to a significant decrease of NO and MDA contents, compared to the control group. Also, CAT and TAA activities were increased significantly. In support of these results, El-Far et al. (2012) demonstrated that injection of MSCs led to a correction and fix of the inequity between ROS and antioxidant protection, thereby enhancing antioxidant-mediated defense and adjusting lipid peroxidation Mohamed et al., 2012.
In the present study, the MSC treated groups were associated with a significant elevation in CAT and TAA activities, compared to G2 and other recovery groups. However, administration of MSCs significantly prohibited the impact of NaNO2 on antioxidative regularity; it reduced NO and MDA and, in parallel, elevated the CAT activities in the tissue of the testes. These data are in agreement with Hussein et al. (2015) who demonstrated the antiperoxidative function of MSCs Hussein et al., 2015. The defense mechanism by bone marrow MSCs, in vivo, against oxidative alteration may be due to its free radical hunting potential. Moreover, the precise reactions of MSCs to oxidative damage may play a vital role in control of tissue homeostasis in addition to renewal of organs following oxidative damage Burova et al., 2013Hassan and Alam, 2014.
Histological changes evoked in the testes by NaNO2 administration were mainly alterations of the seminiferous tubules, including vacuolization, lipid accumulation in intertubular space (with gelatinous material in intertubular connective tissues), and diminution of Leydig cells. Other observed histological changes were reduced germ cell layers, and appearance of hyaline materiel in the lumen of seminiferous tubules and interstitial connective tissues. The results from our study are in accordance with the majority of other works that have revealed changes in the human male constituents of fertility by hypoxia, including reduction in sperm count, reduction in sperm mobility and alleviation in testosterone levels after numerous weeks of exposure.
In addition, the studies showed dilated and congested blood vessels, hyaline material in interstitial connective tissues, and absence of Leydig cells. These results are discussed by several studies Barnholt et al., 2006Okumura et al., 2003. Earlier, Semenza, (2001) which have reported that the increase of vascular endothelial growth factor (VEGF) contents and the representation of their receptors in hypoxic cells are arbitrated by hypoxia inducing factor 1 (HIF-1) Semenza, 2001. Additionally, Hwang et al. (2007) found that VEGF has an effect on the propagation of Leydig cells and on testosterone levels in a dose-dependent manner Hwang et al., 2007. Environmental hypoxia induces vascular alterations that are related with a rise in the temperature of the testes Farias et al., 2005Farías et al., 2008. Recently, Velickovic et al. (2012) reported that at the tissue level, a hypoxic condition induces neovascularization that needs a sequence of actions involving differentiation, proliferation and migration of endothelial cells, as well as formation and maturation of vessels. All these are induced through VEGF Velickovic et al., 2012.
In the present study, most seminiferous tubules showed disorganization of spermatogenic cells, injured germ cells exiting the basal lamina (with vacuoles in between the spermatogenic cells), absence of Sertoli cells, and reduction in number of sperms with hyaline material in between. Our histopathological results are in agreement with observations of Liao et al. (2010) Liao et al., 2010, who found that hypobaric hypoxia animal models show a reduction in germ cell numbers, extremely vacuolated Sertoli cells, pyknotic germ cells, decrease in Leydig cell number, increase of testicular blood vessels diameter, and alterations in testosterone levels. Degeneration and necrosis of some spermatogenic cells in association with other pathological changes were detected in the testicular tissues of rats whose diet is supplemented with variable concentrations of NaNO2. These findings could be due to a defect in the function of Sertoli cells, as reported previously by Grant and Butler (1989) Grant and Butler, 1989. Turner and Lysiak (2008) found that experimental testicular ischemia/reperfusion (IR) in rats and mice caused a reduction in germ cells, seminiferous epithelium vacuolization, alleviations in sperm creation, and apoptosis of germ cells Turner and Lysiak, 2008. The lesions are similar to the results observed in our study.
El-Wakf et al. (2009) El-Wakf et al., 2009 and Ansari et al. (2015) Ansari et al., 2015 reported that NaNO2 induces protein and lipid oxidation, membrane damage, osmotic fragility, and morphological alterations in human erythrocytes. Their data suggested the creation of ROS and resultant production of oxidative stress. The method by which stable reduction in oxygen levels supply or create injury/death of germ cells is likely a rise in intra-testicular ROS. Although ROS possess a physiological function in the spermatogenic procedure, a pathological rise in ROS numbers would negatively impact the endurance and development of germ cells Ramalho-Santos et al., 2008. The toxic consequences of using NaNO2 are believed to be arbitrated by the formation of oxidative ions. This is reinforced by the explanation that NaNO2 raises lipid peroxidation, and lowers GSH levels and protein oxidation in treated tissues Vossen and De Smet, 2015. Moreover, NaNO2 toxicity can be alleviated by antioxidant supplementation Al-Gayyar et al., 2014Al-Gayyar et al., 2016. Pavlova et al., (2013) found that sperm count reduction was observed in all NaNO2 administration experimental groups Pavlova et al., 2013; these alterations could be signs of damaged spermatogenesis.
In the present work, biochemical analyses of the testes tissue revealed that NaNO2 toxicity induced a significant elevation of MDA, NO and DNA F%. Lipid peroxidation has been recommended as one of the basic molecular mechanisms for degeneration and necrosis of some of the spermatogenic cells. In addition, other pathological changes were detected in the testicular tissues of rats whose diet was supplemented with variable concentrations of sodium nitrite Vossen and De Smet, 2015. These findings could be due to defects in the function of Sertoli cells, as reported previously by Grant and Butler (1989) Grant and Butler, 1989. Additionally, formation of reactive nitrogen species by NaNO2 plays a vital role in its carcinogenic effect on cells or different body tissues- activating lipid peroxidation, enzyme inactivation, DNA abrasions and organ injury El-Wakf et al., 2009. These mechanisms could explain the detection of different pathological changes in various organs examined in the present work.
Lim et al. (2014) and Marx et al. (2015) also reported that MSCs are believed to be the main promising cell platform for curative applications Lim et al., 2014Marx et al., 2015. Nevertheless, the proliferation of MSCs and differentiation capability may vary among species and tissue origin. Numerous studies have showed that O2 supply might deeply affect stem cell viability and can support certain types of stem differentiation, while preventing other types of differentiation Simon and Keith, 2008. Spermatogenesis in mammals is a firmly synchronized and incessant process where spermatogonial cells upgrade to the final form of spermatozoa. A physiological hypoxic state keeps self-renewal of spermatogonial cells and spermatogenesis. The vulnerability of the testes to pathological hypo-oxygenic state, particularly chronic hypoxia, is a cause of some types of male infertility Velickovic and Stefanovic, 2014.
Our histological evaluation of rat testes sections of groups treated with MSCs (G3 and G4 groups) showed an improvement of the testicular structure and a relatively normal order of the different stages of spermatogonia. These results are in agreement with Cakici et al. (2013) Cakici et al., 2013, who recorded that MSCs were present in both the external basal partitions and in the internal side of seminiferous tubules. Such observations suggest that MSCs might possess a role in recovering spermatogenesis via two mechanisms: 1) MSC transformation into sperm, or 2) preservation of the spermatogonial cells. Hassan and Alam (2014) showed that a functional rich source for treating infertility is through MSCs Hassan and Alam, 2014. Yazawa et al. (2006) confirmed that MSCs possess the ability to distinguish into steroidogenic cells, like Leydig cells, in vivo and in vitro Yazawa et al., 2006. Lue et al. (2007) revealed that MSCs, injected into testes of infertility busulfan-treated mice, can differentiate into germ cells, Leydig cells and Sertoli cells Lue et al., 2007.
Conclusion
Treatment with NaNO2 induced hypoxia in the rat testes. NaNO2 led to an increase of NO, MDA contents and DNA F% in testes tissues. In addition, a significant decrease was shown in CAT and TAA activities of testes tissues when compared with control rats. Moreover, the histological hypoxic testes sections showed detachment of basement membrane from spermatogenic layers and widening of seminiferous lumen, faintly stained spermatogenic cell layers in different stages (due to vacuolated cytoplasm), and hydropic degeneration and swing shape of sperm cells. Importantly, treatment of rats with MSCs improves toxicity associated with NaNO2 induced hypoxia in the testes of the rats. Overall, the data indicate that MSC therapy can limit damage from hypoxia induced by NaNO2.
Abbreviation
CAT: Catalse
DNA F%: DNA fragmentation percentage
MDA Malondialdhyde
MSCs mesenchymal Stem Cells
NaNO2 Sodium nitrite
NO Nitric oxide
TAA Total antioxidant activity
Author contribution
Prof. Dr Nadia Ismaeil and Dr Amany Osman shared in designing the manuscript and photo and reviewed the histological studies. Prof. Dr. Laila Rashed prepared and suplemented us with the stem cells. Dr. Elham Ali shared in designing the manuscript, interpret the data and reviewed the physiological studies, and Manal Saleh preform the experiment, interpret the data, and write the manuscript.
References
-
M.M.
Al-Gayyar,
A.
Al Youssef,
I.O.
Sherif,
M.E.
Shams,
A.
Abbas.
Protective effects of arjunolic acid against cardiac toxicity induced by oral sodium nitrite: effects on cytokine balance and apoptosis. Life sciences.
2014;
111
:
18-26
.
-
M.M.
Al-Gayyar,
H.M.
Hassan,
A.
Alyoussef,
A.
Abbas,
M.M.
Darweish,
A.A.
El-Hawwary.
Nigella sativa oil attenuates chronic nephrotoxicity induced by oral sodium nitrite: Effects on tissue fibrosis and apoptosis. Redox Report.
2016;
21
:
50-60
.
-
A.
Alhadlaq,
J.J.
Mao.
Mesenchymal stem cells: isolation and therapeutics. Stem cells and development.
2004;
13
:
436-448
.
-
J.G.
Alvarez,
J.C.
Touchstone,
L.
Blasco,
B.T.
Storey.
Spontaneous lipid peroxidation and production of hydrogen peroxide and superoxide in human spermatozoa. Superoxide dismutase as major enzyme protectant against oxygen toxicity. J androl.
1987;
8
:
338-348
.
-
F.A.
Ansari,
S.N.
Ali,
R.
Mahmood.
Sodium nitrite-induced oxidative stress causes membrane damage, protein oxidation, lipid peroxidation and alters major metabolic pathways in human erythrocytes. Toxicology in Vitro.
2015;
29
:
1878-1886
.
-
K.E.
Barnholt,
A.R.
Hoffman,
P.B.
Rock,
S.R.
Muza,
C.S.
Fulco,
B.
Braun,
L.
Holloway,
R.S.
Mazzeo,
A.
Cymerman,
A.L.
Friedlander.
Endocrine responses to acute and chronic high-altitude exposure (4,300 meters): modulating effects of caloric restriction. American Journal of Physiology-Endocrinology and Metabolism.
2006;
290
:
E1078-E1088
.
-
M.
Bhanumathy,
M.
Harish,
H.
Shivaprasad,
G.
Sushma.
Nootropic activity of Celastrus paniculatus seed. Pharmaceutical biology.
2010;
48
:
324-327
.
-
P.
Bock,
R.
Kramer,
M.
Pavelka.
Peroxisomes and related particles. Cell biology monographs.
1980;
7
:
44-74
.
-
S.P.
Bruder,
N.
Jaiswal,
S.E.
Haynesworth.
Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. Journal of cellular biochemistry.
1997;
64
:
278-294
.
-
E.
Burova,
A.
Borodkina,
A.
Shatrova,
N.
Nikolsky.
Sublethal oxidative stress induces the premature senescence of human mesenchymal stem cells derived from endometrium. Oxidative medicine and cellular longevity 2013.
2013
.
-
C.
Cakici,
B.
Buyrukcu,
G.
Duruksu,
A.H.
Haliloglu,
A.
Aksoy,
A.
Isık,
O.
Uludag,
H.
Ustun,
C.
Subası,
E.
Karaoz.
Recovery of fertility in azoospermia rats after injection of adipose -tissue-derived mesenchymal stem cells: the sperm generation. BioMed research international 2013.
2013
.
-
E.J.
Caterson,
L.J.
Nesti,
K.G.
Danielson,
R.S.
Tuan.
Human marrow-derived mesenchymal progenitor cells. Molecular biotechnology.
2002;
20
:
245-256
.
-
M.
Chopp,
X.H.
Zhang,
Y.
Li,
L.
Wang,
J.
Chen,
D.
Lu,
M.
Lu,
M.
Rosenblum.
Spinal cord injury in rat: treatment with bone marrow stromal cell transplantation. Neuroreport.
2000;
11
:
3001-3005
.
-
K.
Cosby,
K.S.
Partovi,
J.H.
Crawford,
R.P.
Patel,
C.D.
Reiter,
S.
Martyr,
B.K.
Yang,
M.A.
Waclawiw,
G.
Zalos,
X.
Xu.
Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nature medicine.
2003;
9
:
1498-1505
.
-
J.C.
David,
J.F.
Grongnet.
Effect of hypoxia on DNA fragmentation in different brain regions of the newborn piglet. Molecular reproduction and development.
2000;
57
:
153-158
.
-
M.
De Ambrogi,
M.
Spinaci,
G.
Galeati,
C.
Tamanini.
Viability and DNA fragmentation in differently sorted boar spermatozoa. Theriogenology.
2006;
66
:
1994-2000
.
-
G.
Dobretsov,
T.
Borschevskaya,
V.
Petrov,
Y.A.
Vladimirov.
The increase of phospholipid bilayer rigidity after lipid peroxidation. FEBS letters.
1977;
84
:
125-128
.
-
H.
Draper,
M.
Hadley.
[43] Malondialdehyde determination as index of lipid peroxidation. Methods in enzymology.
1990;
186
:
421-431
.
-
R.
Drury,
E.
Wallington.
Preparation and fixation of tissues. Carleton's histological technique.
1980;
5
:
41-54
.
-
N.K.
Duru,
M.
Morshedi,
S.
Oehninger.
Effects of hydrogen peroxide on DNA and plasma membrane integrity of human spermatozoa. Fertility and sterility.
2000;
74
:
1200-1207
.
-
M.F.
El Asmar,
H.M.
Atta,
S.
Mahfouz,
H.H.
Fouad,
N.K.
Roshdy,
L.A.
Rashed,
D.
Sabry,
A.A.
Hassouna,
F.M.
Taha.
Efficacy of mesenchymal stem cells in suppression of hepatocarcinorigenesis in rats: possible role of Wnt signaling. Journal of Experimental & Clinical Cancer Research.
2011;
30
:
49
.
-
N.M.
El-Sheikh,
F.A.
Khalil.
L-Arginine and L-glutamine as immunonutrients and modulating agents for oxidative stress and toxicity induced by sodium nitrite in rats. Food and Chemical Toxicology.
2011;
49
:
758-762
.
-
A.
El-Wakf,
H.
Hassan,
F.
El-Said,
A.
El-Said.
The association between nitrite contamination of drinking water and changes in methemoglobin level other hematological parameters in young and adult male rats J. Egypt Soc Toxicol.
2009;
49
:
91-96
.
-
J.
Farias,
E.
Bustos-Obregón,
R.
Orellana,
J.
Bucarey,
E.
Quiroz,
J.
Reyes.
Effects of chronic hypobaric hypoxia on testis histology and round spermatid oxidative metabolism. Andrologia.
2005;
37
:
47-52
.
-
J.G.
Farías,
E.
Bustos-Obregon,
P.J.
Tapia,
E.
Gutiérrez,
A.
Zepeda,
C.
Juantok,
C.
Gonzalo,
S.
Gustavo,
J.
Benites,
J.G.
Reyes.
Time course of endocrine changes in the hypophysis-gonad axis induced by hypobaric hypoxia in male rats. Journal of Reproduction and Development.
2008;
54
:
18-21
.
-
Y.
Fujimoto,
M.
Abematsu,
A.
Falk,
K.
Tsujimura,
T.
Sanosaka,
B.
Juliandi,
K.
Semi,
M.
Namihira,
S.
Komiya,
A.
Smith.
Treatment of a mouse model of spinal cord injury by transplantation of human induced pluripotent stem cell-derived long-term self-renewing neuroepithelial-like stem cells. Stem Cells.
2012;
30
:
1163-1173
.
-
M.
Georgiou,
L.M.
Perkins,
A.H.
Payne.
Steroid synthesis-dependent, oxygen-mediated damage of mitochondrial and microsomal cytochrome P-450 enzymes in rat Leydig cell cultures. Endocrinology.
1987;
121
:
1390-1399
.
-
D.
Grant,
W.
Butler.
Chronic toxicity of sodium nitrite in the male F344 rat. Food and Chemical Toxicology.
1989;
27
:
565-571
.
-
P.
Hall.
Testicular steroid synthesis: organization and regulation. The physiology of reproduction.
1994;
1
:
1335-1362
.
-
A.I.
Hassan,
S.S.
Alam.
Evaluation of mesenchymal stem cells in treatment of infertility in male rats. Stem cell research & therapy.
2014;
5
:
131
.
-
H.A.
Hassan,
S.M.
El-Agmy,
R.L.
Gaur,
A.
Fernando,
M.
Raj,
A.
Ouhtit.
In vivo evidence of hepato-and reno-protective effect of garlic oil against sodium nitrite-induced oxidative stress. Int J Biol Sci.
2009;
5
:
249-255
.
-
Y.
Hussein,
R.
Hussein,
A.
Amin,
H.
Hussein.
Evaluation of Mesenchymal Stem Cells and Vitamin E in Treatment of Infertile Male Albino Rats. Int J of Multidisciplinary and Current research.
2015;
3
.
-
G.-S.
Hwang,
S.-W.
Wang,
W.-M.
Tseng,
C.-H.
Yu,
P.S.
Wang.
Effect of hypoxia on the release of vascular endothelial growth factor and testosterone in mouse TM3 Leydig cells. American Journal of Physiology-Endocrinology and Metabolism.
2007;
292
:
E1763-E1769
.
-
T.
Ishikawa,
Y.
Kondo,
K.
Goda,
M.
Fujisawa.
Overexpression of endothelial nitric oxide synthase in transgenic mice accelerates testicular germ cell apoptosis induced by experimental cryptorchidism. Journal of andrology.
2005;
26
:
281-288
.
-
N.
Jaiswal,
S.E.
Haynesworth,
A.I.
Caplan,
S.P.
Bruder.
Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. Journal of cellular biochemistry.
1997;
64
:
295-312
.
-
P.
Kebriaei,
L.
Isola,
E.
Bahceci,
K.
Holland,
S.
Rowley,
J.
McGuirk,
M.
Devetten,
J.
Jansen,
R.
Herzig,
M.
Schuster.
Adult human mesenchymal stem cells added to corticosteroid therapy for the treatment of acute graft-versus-host disease. Biology of Blood and Marrow Transplantation.
2009;
15
:
804-811
.
-
A.M.
Knaapen,
R.P.
Schins,
P.J.
Borm,
F.J.
van Schooten.
Nitrite enhances neutrophil-induced DNA strand breakage in pulmonary epithelial cells by inhibition of myeloperoxidase. Carcinogenesis.
2005;
26
:
1642-1648
.
-
H.
Kodama,
Y.
Kuribayashi,
C.
Gagnon.
Effect of sperm lipid peroxidation on fertilization. Journal of andrology.
1996;
17
:
151-157
.
-
D.
Koracevic,
G.
Koracevic,
V.
Djordjevic,
S.
Andrejevic,
V.
Cosic.
Method for the measurement of antioxidant activity in human fluids. Journal of clinical pathology.
2001;
54
:
356-361
.
-
R.-M.
Laberge,
G.
Boissonneault.
Chromatin remodeling in spermatids: a sensitive step for the genetic integrity of the male gamete. Archives of andrology.
2005;
51
:
125-133
.
-
W.
Liao,
M.
Cai,
J.
Chen,
J.
Huang,
F.
Liu,
C.
Jiang,
Y.
Gao.
Hypobaric hypoxia causes deleterious effects on spermatogenesis in rats. Reproduction.
2010;
139
:
1031-1038
.
-
M.H.
Lim,
W.K.
Ong,
S.
Sugii.
The current landscape of adipose-derived stem cells in clinical applications. Expert reviews in molecular medicine.
2014;
16
:
e8
.
-
Y.
Lue,
K.
Erkkila,
P.Y.
Liu,
K.
Ma,
C.
Wang,
A.S.
Hikim,
R.S.
Swerdloff.
Fate of bone marrow stem cells transplanted into the testis: potential implication for men with testicular failure. The American journal of pathology.
2007;
170
:
899-908
.
-
A.K.
Mack,
I.
McGowan,
R.
Vicki,
C.K.
Tremonti,
D.
Ackah,
C.
Barnett,
R.F.
Machado,
M.T.
Gladwin,
G.J.
Kato.
Sodium nitrite promotes regional blood flow in patients with sickle cell disease: a phase I/II study. British journal of haematology.
2008;
142
:
971-978
.
-
A.R.
Maher,
A.B.
Milsom,
P.
Gunaruwan,
K.
Abozguia,
I.
Ahmed,
R.A.
Weaver,
P.
Thomas,
H.
Ashrafian,
G.V.
Born,
P.E.
James.
Hypoxic modulation of exogenous nitrite-induced vasodilation in humans. Circulation.
2008;
117
:
670-677
.
-
C.
Marx,
M.D.
Silveira,
N.
Beyer Nardi.
Adipose-derived stem cells in veterinary medicine: characterization and therapeutic applications. Stem cells and development.
2015;
24
:
803-813
.
-
K.M.
Miranda,
M.G.
Espey,
D.A.
Wink.
A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric oxide.
2001;
5
:
62-71
.
-
A.
Mohamed,
M.M.
Gabr,
S.M.
El-Halawani,
R.Y.
Ibrahim,
S.M.
Khater,
A.F.
Refaie,
M.A.
Ghoneim.
Novel evidence of restoring and augmenting antioxidant defense after treatment of diabetic rats using stem cells. Curr Top Biochem Res.
2012;
14
:
25-37
.
-
M.H.
Moustafa,
R.K.
Sharma,
J.
Thornton,
E.
Mascha,
M.A.
Abdel-Hafez,
A.J.
Thomas,
A.
Agarwal.
Relationship between ROS production, apoptosis and DNA denaturation in spermatozoa from patients examined for infertility. Human Reproduction.
2004;
19
:
129-138
.
-
K.
Nayernia,
J.
Nolte,
H.W.
Michelmann,
J.H.
Lee,
K.
Rathsack,
N.
Drusenheimer,
A.
Dev,
G.
Wulf,
I.E.
Ehrmann,
D.J.
Elliott.
In vitro-differentiated embryonic stem cells give rise to male gametes that can generate offspring mice. Developmental cell.
2006;
11
:
125-132
.
-
U.
Nöth,
A.M.
Osyczka,
R.
Tuli,
N.J.
Hickok,
K.G.
Danielson,
R.S.
Tuan.
Multilineage mesenchymal differentiation potential of human trabecular bone-derived cells. Journal of Orthopaedic Research.
2002;
20
:
1060-1069
.
-
A.
Okumura,
H.
Fuse,
Y.
Kawauchi,
I.
Mizuno,
T.
Akashi.
Changes in male reproductive function after high altitude mountaineering. High altitude medicine & biology.
2003;
4
:
349-353
.
-
R.
Oliva.
Protamines and male infertility. Human reproduction update.
2006;
12
:
417-435
.
-
A.
Özdamar,
A.
Soylu,
M.
Culha,
M.
Özden,
A.
Gökalp.
Testicular oxidative stress. Urologia internationalis.
2004;
73
:
343-347
.
-
E.
Pavlova,
D.
Dimova,
E.
Petrova,
Y.
Gluhcheva,
N.
Atanassova.
Changes in rat testis and sperm count after acute treatment with sodium nitrite. Bulg J Agric Sci.
2013;
19
:
186-189
.
-
M.F.
Pittenger,
A.M.
Mackay,
S.C.
Beck,
R.K.
Jaiswal,
R.
Douglas,
J.D.
Mosca,
M.A.
Moorman,
D.W.
Simonetti,
S.
Craig,
D.R.
Marshak.
Multilineage potential of adult human mesenchymal stem cells. science.
1999;
284
:
143-147
.
-
J.
Ramalho-Santos,
S.
Amaral,
P.J.
Oliveira.
Diabetes and the impairment of reproductive function: possible role of mitochondria and reactive oxygen species. Current diabetes reviews.
2008;
4
:
46-54
.
-
C.
Romeo,
R.
Ientile,
P.
Impellizzeri,
N.
Turiaco,
M.
Teletta,
P.
Antonuccio,
M.
Basile,
C.
Gentile.
Preliminary report on nitric oxide-mediated oxidative damage in adolescent varicocele. Human Reproduction.
2003;
18
:
26-29
.
-
M.F.
Salama,
A.
Abbas,
M.M.
Darweish,
A.A.
El-Hawwary,
M.M.
Al-Gayyar.
Hepatoprotective effects of cod liver oil against sodium nitrite toxicity in rats. Pharmaceutical biology.
2013;
51
:
1435-1443
.
-
Y.
Sato,
H.
Araki,
J.
Kato,
K.
Nakamura,
Y.
Kawano,
M.
Kobune,
T.
Sato,
K.
Miyanishi,
T.
Takayama,
M.
Takahashi.
Human mesenchymal stem cells xenografted directly to rat liver are differentiated into human hepatocytes without fusion. Blood.
2005;
106
:
756-763
.
-
G.L.
Semenza.
Hypoxia-inducible factor 1: control of oxygen homeostasis in health and disease. Pediatric research.
2001;
49
:
614-617
.
-
M.-S.
Seo,
Y.-H.
Jeong,
J.-R.
Park,
S.-B.
Park,
K.-H.
Rho,
H.-S.
Kim,
K.-R.
Yu,
S.-H.
Lee,
J.-W.
Jung,
Y.-S.
Lee.
Isolation and characterization of canine umbilical cord blood-derived mesenchymal stem cells. Journal of veterinary science.
2009;
10
:
181-187
.
-
F.K.T.
Sharawy.
Experimental studies on the effect of the antifungal drug itraconazole on pregnant rats and their fetuses. In Faculty of Science (Cairo University).
2013
.
-
M.C.
Simon,
B.
Keith.
The role of oxygen availability in embryonic development and stem cell function. Nature reviews Molecular cell biology.
2008;
9
:
285-296
.
-
V.Y.
Titov,
Y.M.
Petrenko.
Nitrite-catalase interaction as an important element of nitrite toxicity. Biochemistry (Moscow).
2003;
68
:
627-633
.
-
T.T.
Turner,
J.J.
Lysiak.
Oxidative stress: a common factor in testicular dysfunction. Journal of Andrology.
2008;
29
:
488-498
.
-
DHHS
U.S..
Department of Health and Human Services. Toxicology and Carcinogenesis Studies of Sodium Nitrite. Drinking Water Studies.
2001
.
-
Á.
Vargas,
E.
Bustos-Obregón,
R.
Hartley.
Effects of hypoxia on epididymal sperm parameters and protective role of ibuprofen and melatonin. Biological research.
2011;
44
:
161-167
.
-
L.J.
Velickovic,
A.R.
Petrovic,
S.
Stojnev,
Z.
Dolicanin,
T.
Hattori,
H.
Sugihara,
K.-i.
Mukaisho,
M.
Stojanovic,
V.
Stefanovic.
Angiogenesis in upper tract urothelial carcinoma associated with Balkan endemic nephropathy. International journal of clinical and experimental pathology.
2012;
5
:
674
.
-
L.J.
Velickovic,
V.
Stefanovic.
Hypoxia and spermatogenesis. International urology and nephrology.
2014;
46
:
887-894
.
-
J.
Villegas,
M.
Schulz,
L.
Soto,
T.
Iglesias,
W.
Miska,
R.
Sánchez.
Influence of reactive oxygen species produced by activated leukocytes at the level of apoptosis in mature human spermatozoa. Fertility and sterility.
2005;
83
:
808-810
.
-
E.
Vossen,
S.
De Smet.
Protein oxidation and protein nitration influenced by sodium nitrite in two different meat model systems. Journal of agricultural and food chemistry.
2015;
63
:
2550-2556
.
-
X.
Wang,
R.K.
Sharma,
S.C.
Sikka,
A.J.
Thomas,
T.
Falcone,
A.
Agarwal.
Oxidative stress is associated with increased apoptosis leading to spermatozoa DNA damage in patients with male factor infertility. Fertility and sterility.
2003;
80
:
531-535
.
-
T.
Yazawa,
T.
Mizutani,
K.
Yamada,
H.
Kawata,
T.
Sekiguchi,
M.
Yoshino,
T.
Kajitani,
Z.
Shou,
A.
Umezawa,
K.
Miyamoto.
Differentiation of adult stem cells derived from bone marrow stroma into Leydig or adrenocortical cells. Endocrinology.
2006;
147
:
4104-4111
.
Comments
Downloads
Article Details
Volume & Issue : Vol 4 No 02 (2017)
Page No.: 1128-1146
Published on: 2017-02-20
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 6261 times
- Download PDF downloaded - 2333 times
- View Article downloaded - 34 times

{kind=link}
{kind=link}
{kind=link}